Eukaryotic
cells are dependent on
oxygen levels as well as the
presence of
functional mitochondria in order to efficiently generate ATP through
oxidative phosphorylation (OXPHOS). Cells respond quickly to changes in
oxygen levels by activating several signaling pathways that provide
metabolic and adaptive mechanisms to the new environment.
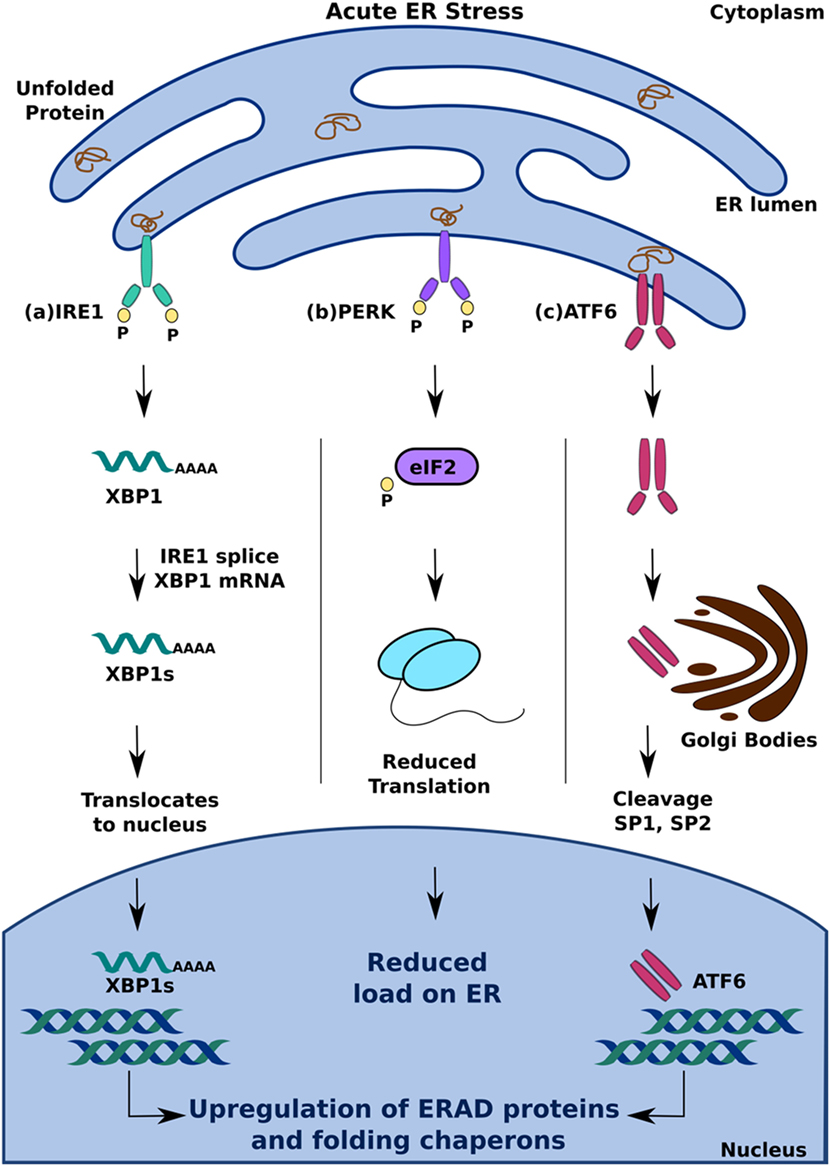
Hypoxia-inducible factor 1α (HIF-1α) is the primary transcriptional
regulator of the
cell response to low oxygen levels (hypoxia)1,2,3,4.
Accumulation of HIF-1α protein and
its translocation to the
cell
nucleus leads to the transcriptional
activation of several hundred genes
that carry a hypoxia response element (HRE) in their promoters
5,6.
This leads to HIF-1α-dependent
reprogramming of cellular metabolism
that shifts the ATP production from oxidative phosphorylation, that is
limited under low oxygen levels, to
glycolysis7,8.
There is an important phenomenon associated with
most cancer cells
where glycolysis is predominantly used as the main source of ATP
production, even under normal levels of oxygen (normoxia). This is known
as the Warburg effect, originally described in 1923 and has since been
extensively studied
9,10.
Accumulating evidence indicates that induced
aerobic glycolysis is not
only the hallmark of cancer, but it is also important in many cellular
processes including embryogenesis, innate and adaptive immunity, type 2
diabetes, starvation, as well as cardiomyopathy
11,12,13,14,15,16,17.
The mechanism that potentially “bypasses” the tight regulation of
cellular metabolism by the HIF-1α transcription factor is unknown.
Under
normoxia, HIF-1α is continuously synthesized and
degraded in the
cytosol through a highly regulated process. The
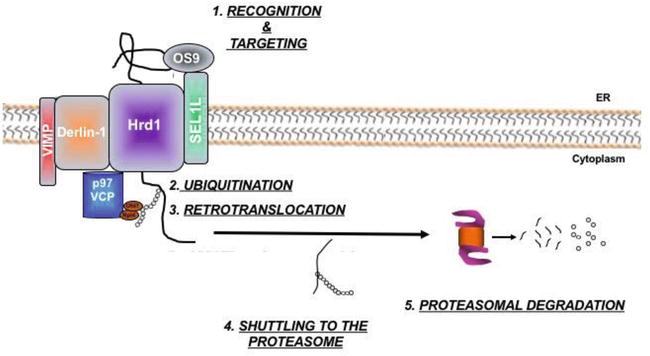
oxygen sensor propyl
hydroxylase 2 (PHD2) hydroxylates HIF-1α which then binds the von
Hippel-Lindau (VHL) tumor suppressor protein and gets ubiquitinated by
the CRL2
VHL ligase complex
18.
We
have uncovered
a new pathway that regulates HIF-1α protein levels and
involves the
mitochondrial MUL1 E3 ubiquitin ligase. MUL1 (also known as
Mulan, MAPL, GIDE, and HADES) is
one of three mitochondrial E3
ubiquitin ligases, the other two being MARCH5 and RNF185
19,20,21,22,23,24,25,26,27,28.
Previous studies have shown MUL1 to be a major player in a number of
pathways involved in apoptosis, mitophagy, and innate immune response
21,25,26,29,30,31,32,33. MUL1 is able to modify specific substrates through SUMOylation, as well as K63- or K48-ubiquitination
20,21,34,35.
Our data show that MUL1, through K48-ubiquitination, directly regulates
the level of UBXN7 protein, also known as UBX domain protein 7 (UBXD7)
36.
UBXN7 serves as a substrate binding adaptor and interacts with several
proteins including HIF-1α, CUL2, as well as AAA + ATPase p97, also known
as VCP (Valosin-containing protein)
36,37.
We identified lysine 14 (K14) and lysine 412 (K412) on the UBXN7
protein as the two major K48-ubiquitination sites for MUL1 ligase.
Ubiquitination of UBXN7 targets the protein for proteasome degradation
and inactivation of MUL1 leads to high levels of UBXN7 with concomitant
increase in HIF-1α protein. The accumulation of HIF-1α is functional and
is accompanied by activation of GLUT1, a known target of HIF-1α
38.
This deregulation of HIF-1α affects the metabolic state of cells with
glycolysis becoming the predominant energy production pathway even
during aerobic conditions. In summary, we describe a new
mitochondrial-initiated pathway that interferes with the process of
HIF-1α regulation and reprograms cellular metabolism to induce aerobic
glycolysis
Results
Schematic diagram of the proposed new pathway that operates upstream of the CRL2VHL
complex and involves the UBXN7 cofactor protein and its regulation by
mitochondrial MUL1 E3 ligase. MUL1 ligase, through constant K48-linked
polyubiquitination, maintains a steady low level of UBXN7 protein that
is able to act as cofactor in assembling the active CRL2VHL
complex necessary for the regulation of HIF-1α during normoxia. When
MUL1 becomes inactive or its activity is compromised, it leads to high
levels of UBXN7 protein that function as an inhibitor of the CRL2VHL complex. Without an active CRL2VHL
complex HIF-1α protein accumulates and drives glycolysis under
normoxia. MUL1 protein levels are regulated by K48-autoubiquitination as
well as by the action of the mitochondrial Omi/HtrA2 protease.
---------
This is the first report to show the existence of a new pathway where
mitochondria, through MUL1, regulate HIF-1α protein levels. Our data
clearly show this pathway is very important under normal conditions and
could potentially be involved in the mitochondrial induction of aerobic
glycolysis.
 a,b,c,d
a,b,c,d {kind=link}
{kind=link}
{kind=link}