HIV-1 integrase reveals critical regions for protein–DNA interaction
Dominic Esposito1 and Robert Craigie1
- Laboratory of Molecular Biology, NIDDK, National Institutes of Health, 5 Center Drive MSC0560, Bethesda, MD 20892, USA
Correspondence to:
Robert Craigie, E-mail: bobc@helix.nih.gov
Received 6 July 1998; Accepted 5 August 1998; Revised 29 July 1998
Abstract (Tiivistelmä)
- HIV-1 integraasi tunnistaa ja pilkkoo viruksen DNA:n päädyn alkuaskelena integaatioprosessiin.
HIV-1 integrase specifically recognizes and cleaves viral end DNA during the initial step of retroviral integration.
- Mitkä seikat proteiinissa ja DNA:ssa määräävät tämän spesifisen sitoutumisen viruksen päättyyn?
The protein and DNA determinants of the specificity of viral end DNA binding have not been clearly identified.
- Asiaa tutkitaan: Analuysoidaan viruksen DNApäädyn LTR-sekvenssi ja sen interaktiot integraasiin.
We have used mutational analysis of the viral end LTR sequence, in vitro selection of optimal viral end sequences, and specific photocrosslinking to identify regions of integrase that interact with specific bases in the LTR termini.
- Integraasin CCD jakson ( integraasin ydindomaanin) epäjärjestäytynyt silmukka erityisesti aminohapot Q148 ja Y143 osallistuvat sitoutumalla viruksen DNA:n päätyyn.
The results highlight the involvement of the disordered loop of the integrase core domain, specifically residues Q148 and Y143, in binding to the terminal portion of the viral DNA ends.
- Lisäksi löytyy v ds DNA.ssa ylävirtaan sijaitsevana LTR-päädyssä integraasin C-terminaalisen domaanin kanssa interaktiossa olevia asemia jotka stabiloivat viruksen DNA:n sitoutumista.
Additionally, we have identified positions upstream in the LTR termini which interact with the C-terminal domain of integrase, providing evidence for the role of that domain in stabilization of viral DNA binding.
- Lisäksi havaitaan 12 emästä DNA-päädystä laskien sijaitseva alue, joka on essentielli DNA:n sitoutumisessa magnesiumin (Mg++) läsnäollessa mutta ei manganeesin (Mn++) läsnäollsessa, viitaten eri divalenttien kationien merkityksesn sekvenssispesifisessä sitoutumisessa.
Finally, we have located a region centered 12 bases from the viral DNA terminus which appears essential for viral end DNA binding in the presence of magnesium, but not in the presence of manganese, suggesting a differential effect of divalent cations on sequence-specific binding.
- Nämäkin tulokset auttavat määrittämään tärkeitä alueita integraasin(IN) ja virusDNA: n välisessä kontaktissa ja avustavat hahmottamaan tämän vitaalin interaktion molekulaarista kaavaa.
These results help to define important regions of contact between integrase and viral DNA, and assist in the formulation of a molecular model of this vital interaction.
Paul Spearmanin kirjassa kerrotaan tarkasti eri retrovirusten tärkeistä sekvenseistä kuten HIV-1 integraasista seuraavaa sivulla 134:
Aminohapot 90- 190 ovat seuraavia90 PAETG QETAYF ( osaa alfa1helixiä)
100 LLKL AGRWPV (osa alfa-1 helixiä)
110 KTVH T DNGSN
120 FTSTT VKAAS ( alfa2 helixiä)(osaa alfa3 helixiä)
130 WWAGI KQEFGI ( osaa alfa 3-helixiä)
140 PYNPQ SQGVI (osaa alfa4 helixiä)
150 ESMNK ELKKI(alfa4 helixiä)
160IGQVR DQAEH (osaa alfa 4 helixiä) (alfa 4/5 connectorpeptidi)
170 LKTAV QMAVF (osaa alfa 5 helixiä)
180 IHNFKRKGG (osaa alfa 5 helixiä)
190
Kaikilla viruksilla HIV, SIV, FIV, BIV, EIAV on alfa1-helixissä samassa asemassa leusiini(L).
Samoin aspasrtaatti (D) alfahelixien 1 ja 2 välillä. ( Tämän jälkeen on yksi katalyyttinen keskus N)
Alfa3 sekvenssi on kaikilla melko erilainen: HIV-1 viruksella: TVKAASWWA
Aminohappo 150E on taas kaikissa sama. (Tämän jälkeen on toinen katalyyttinen keskus M)
Alfa4/5 konnektoripeptidi on suhteellisen erilainen kaikilla.
HIV-1 alfa4/5 konnektoripeptidi on jaksolla DQAEHLKTAVQMA
Katalyyttiset keskukset eivät käy kontaktiin IBD-pinnan kanssa.
Integraasin pitää ainakin dimerisoitua, jotta se voi koeputkessa tehdä 3´päädyn prosessoinnin, trimmauksen. Mutta tetrameeri vaaditaan DNA säikeen siirtoon. Isompiakin multimeerejä on havaittu HIV-2 viruksen yhteydessä. Mikä relevanssi oligomeerisella tilalla on?
Muistettava HAART terapian nykyarsenaali:
Classes of drugs
Antiretroviral (ARV) drugs are broadly classified by the phase of the retrovirus life-cycle that the drug inhibits.
- Entry inhibitors (or fusion inhibitors) interfere with binding, fusion and entry of HIV-1 to the host cell by blocking one of several targets. Maraviroc and enfuvirtide are the two currently available agents in this class.
- CCR5 receptor antagonists are the first antiretroviral drugs which do not target the virus directly. Instead, they bind to the CCR5 receptor on the surface of the T-Cell and block viral attachment to the cell. Most strains of HIV attach to T-Cells using the CCR5 receptor. If HIV cannot attach to the cell, it cannot gain entry to replicate.
- Non-Nucleoside and nucleotide reverse transcriptase inhibitors (NNRTI) inhibit reverse transcription by being incorporated into the newly synthesized viral DNA strand as a faulty nucleotide. This causes a chemical reaction resulting in DNA chain termination.
- Nucleoside reverse transcriptase inhibitors (NRTI) mimic nucleotides and inhibit reverse transcriptase directly by binding to the enzymes polymerase site and interfering with its function.
- Protease inhibitors (PIs) target viral assembly by inhibiting the activity of protease, an enzyme used by HIV to cleave nascent proteins for the final assembly of new virions.
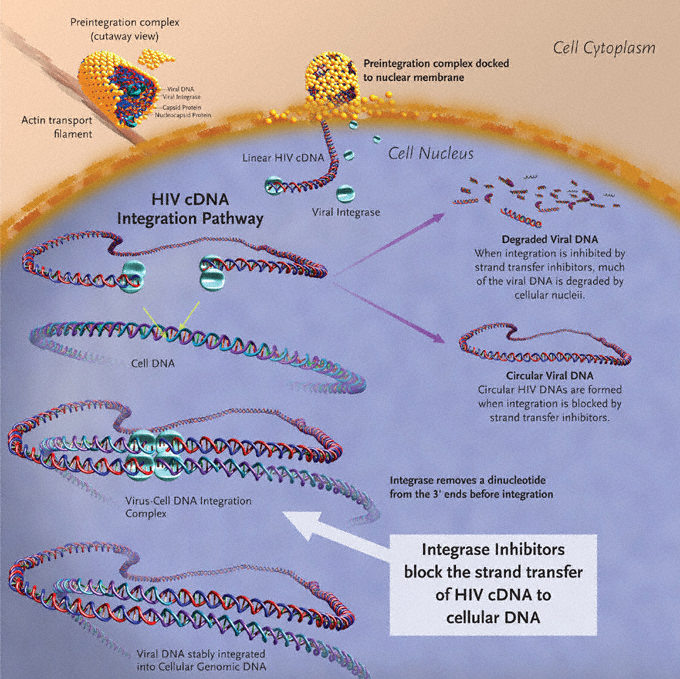
- Integrase inhibitors inhibit the enzyme integrase, which is responsible for integration of viral DNA into the DNA of the infected cell. There are several integrase inhibitors currently under clinical trial, and raltegravir became the first to receive FDA approval in October 2007.
- Maturation inhibitors inhibit the last step in gag processing in which the viral capsid polyprotein is cleaved, thereby blocking the conversion of the polyprotein into the mature capsid protein (p24). Because these viral particles have a defective core, the virions released consist mainly of non-infectious particles. Alpha interferon is a currently available agent in this class.[2] Two additional inhibitors under investigation are bevirimat [3] and Vivecon.


{kind=link}