Molecular Membrane Biology, Department of Cell Biology, Nencki Institute of Experimental Biology of the Polish Academy of Sciences, Warsaw, Poland
S-palmitoylation is a reversible, enzymatic posttranslational modification of proteins in which palmitoyl chain is attached to a cysteine residue via a thioester linkage. S-palmitoylation determines the functioning of proteins by affecting their association with membranes, compartmentalization in membrane domains, trafficking, and stability. In this review, we focus on S-palmitoylation of proteins, which are crucial for the interactions of pathogenic bacteria and viruses with the host. We discuss the role of palmitoylated proteins in the invasion of host cells by bacteria and viruses, and those involved in the host responses to the infection. We highlight recent data on protein S-palmitoylation in pathogens and their hosts obtained owing to the development of methods based on click chemistry and acyl-biotin exchange allowing proteomic analysis of protein lipidation. The role of the palmitoyl moiety present in bacterial lipopolysaccharide and lipoproteins, contributing to infectivity and affecting recognition of bacteria by innate immune receptors, is also discussed.
Introduction
Palmitic acid (C16:0) is a long-chain saturated fatty
acid, and a component of various lipids playing important roles in cell
membrane organization, signal transduction, and energy storage.
Moreover, the palmitoyl chain can be attached to proteins in a process
called palmitoylation (S-palmitoylation), which modification affects their localization and functioning.
In the human body palmitic acid is synthetized in a process called de novo lipogenesis. It takes place mainly in adipocytes, hepatocytes, and cells of lactating mammary glands (1).
Palmitic acid is used for the synthesis of phospholipids and
sphingolipids, may undergo elongation and/or desaturation into other
fatty acids (e.g., stearic acid or oleic acid, respectively), and can be
esterified to form storage lipids—triacylglycerols. Apart from de novo
synthesis, palmitic acid is also provided to the human body with food.
Since palmitic acid is universally found in natural fats, its
consumption exceeds the consumption of other saturated fatty acids and,
in the USA it accounts for about 60% of the total intake of saturated
fatty acids (2).
A growing body of experimental and clinical evidence points to a link
between a westernized diet, including a high intake of saturated fatty
acids, and chronic inflammatory diseases (3–5).
As dietary saturated and unsaturated fatty acids apparently modulate
activity of immune cells, their influence on the immune responses
triggered upon infection is also beginning to be investigated (6).
These facts drive the interest in palmitic acid with an aim of
elucidating the molecular mechanisms of its immunomodulatory properties.
In this review, we focus on S-palmitoylation of
proteins crucial for the interactions of pathogenic bacteria and viruses
with the host. We emphasize novel data on the role of S-palmitoylated
proteins in the invasion of host cells by pathogens and those involved
in the host innate immune responses to the infection, which have been
obtained thanks to the application of new technical approaches.
Recently, substantial progress in the understanding of protein
palmitoylation was made possible by the development of methods allowing
high-throughput analysis of cellular/tissue palmitoyl proteomes. We
begin, however, by showing how unique protein S-palmitoylation is among other protein lipidations.
The Many Faces of Fatty Acylation of Proteins
S-Palmitoylation of Proteins and Its Influence on Protein Localization, Trafficking, and Stability
S-palmitoylation is a posttranslational
modification of proteins consisting in a potentially reversible covalent
attachment of palmitoyl chain to a cysteine residue(s) of proteins
through a thioester bond (Table 1). Thus, S-palmitoylation
resembles other reversible regulatory posttranslational protein
modifications, including phosphorylation or acetylation,
well-established factors affecting protein structure and functions. In
particular, S-palmitoylation modifies cellular localization of
proteins and their stability. The most dramatic changes of localization
concern cytosolic proteins which upon S-palmitoylation acquire a hydrophobic anchor facilitating their docking into membranes (Figure 1). However, several integral membrane proteins also undergo S-palmitoylation.
It often occurs on cysteine residue(s) located in the proximity of the
junction of the transmembrane and cytoplasmic domains of the protein. S-palmitoylated
transmembrane proteins occupy various cellular compartments, such as
endoplasmic reticulum, Golgi apparatus, and the plasma membrane. In
accordance, for some proteins, such as transmembrane adaptor proteins in
leukocytes, S-palmitoylation was found secondary to the length
and hydrophobicity of the transmembrane domain as a determinant of
plasma membrane destination (7).
S-palmitoylation also contributes to the compartmentalization of
proteins to distinct domains of membranes—rafts and tetraspanin-rich
microdomains. In fact, the interest in S-palmitoylation was
boosted when it was found to be required for the targeting of some
signaling proteins to rafts. Rafts are nanodomains of the plasma
membrane and some intracellular membranes, mainly of the trans-Golgi apparatus, rich in sphingolipids, glycerophospholipids with saturated fatty acid chains, and cholesterol (32).
The plasma membrane nanodomains are sites of signal transduction by
distinct receptors of immune cells involved in both acquired immune
reactions, such as T cell receptor (TCR), Fcε receptor I, Fcγ receptor
II, and in innate immune responses, such as toll-like receptor 4 (TLR4) (33, 34).
Rafts are also sites of virion assembly and budding, as established,
e.g., for influenza A virus and human immunodeficiency virus-1 (HIV-1) (35, 36).
Peripheral membrane proteins acylated with saturated fatty acids are
likely to anchor preferentially between the ordered saturated lipids of
rafts rather than between the disordered lipids of the surrounding
membrane. It has been shown that, owing to their raft localization, S-palmitoylated
kinases of the Src family interact with raft-associating plasma
membrane immunoreceptors and initiate signaling cascades fundamental to
acquired immunity (15, 37, 38).
It is worth noting that also the acyl chains attached to proteins can
affect the membrane structure. Studies on model membranes have revealed
that palmitic and myristic acids facilitate formation of ordered
lamellar membrane regions (39, 40). In accordance, S-palmitoylation
of erythrocyte peripheral membrane protein called
membrane-palmitoylated protein 1 (MPP1) was found to be required for the
proper lateral organization and fluidity of erythrocyte membrane. In
the absence of MPP1 S-palmitoylation, raft assembly was disturbed
and erythrocyte functioning compromised leading to hemolytic anemia in
patients deficient in the enzyme catalyzing this reaction (41, 42). Preferential raft association is a feature of some S-palmitoylated
transmembrane proteins, e.g., adaptor proteins PAG, LAT, and NTAL,
which collaborate with the abovementioned immunoreceptors. In fact,
palmitoylation is required for the raft association of most integral
raft proteins (8, 43, 44).
On the other hand, S-palmitoylation does not obligatorily confer raft localization on transmembrane proteins. Certain S-palmitoylated
proteins, such as transferrin receptor, glycoprotein G of vesicular
stomatitis virus (VSV), and anthrax toxin receptor, tumor endothelial
marker 8 (TEM8), are actually excluded from rafts. Apparently, a
combination of S-palmitoylation and the properties of the
transmembrane domain of the protein contribute to its destination to the
raft or non-raft environment (43, 45).
It has also been proposed that the attachment of a fatty acyl chain at
the juxtamembrane cysteine(s) of a protein can induce tilting of its
transmembrane fragment, determining in which part of the membrane it
will accommodate to avoid a hydrophobic mismatch potentially caused by
the thickness of the bilayer (46).
That not all S-palmitoylated proteins associate
with rafts has been shown convincingly for macrophage-like RAW264 cells,
where only about half of those proteins were found in the Triton
X-100-resistant membrane fraction enriched in rafts (47, 48). In accordance, proteomic data on the distribution of S-palmitoylated
proteins in prostate cancer cells have revealed that several such
proteins are recovered in the non-raft (Triton X-100-soluble) fraction
and are likely localized to microdomains enriched in scaffold proteins
called tetraspanins (49).
The tetraspanins are small integral membrane proteins found in the
plasma membrane and other cellular membranes, having four transmembrane
helices and undergoing S-palmitoylation at several conserved
cysteine residues. The tetraspanins interact with each other and with
various transmembrane and cytosolic partners, often also S-palmitoylated, forming microdomains (“tetraspanin web”) (50). It has been suggested that the amino acid composition of the S-palmitoylation
site in some transmembrane proteins, such as the adaptor proteins
involved in acquired immune responses, determines the association of
those S-palmitoylated proteins with rafts or with the tetraspanin-enriched microdomains (44).
An intriguing and still poorly addressed question concerns the relation
between rafts and the tetraspanin-enriched microdomains, apparently of
functional significance, e.g., during virus budding from host cells (35). This uncertainty stems partially from the fact that S-palmitoylation
of tetraspanins governs their interactions with cholesterol and
gangliosides leading at certain conditions to the recovery of
tetraspanins in detergent-resistant membrane fractions enriched in rafts
(51, 52). Besides its involvement in targeting proteins to rafts or tetraspanin-enriched microdomain, S-palmitoylation
has been found to govern accumulation of the transmembrane chaperone
protein calnexin in the perinuclear domain of endoplasmic reticulum (53).
S-palmitoylation also affects protein stability
through its interplay with ubiquitination or phosphorylation, as found
for the anthrax toxin receptor TEM8, antiviral interferon-induced
transmembrane protein IFITM1, calnexin, and zDHHC6, one of palmitoyl
acyltransferases described below (54–57).
- ZDHHC zincfinger proteiinit https://www.genenames.org/data/genegroup/#!/group/76
Also known as : "zf-DHHC", "DHHC palmitoyltransferase domain containing", "Protein acyl transferases", "PATs", "Palmitoyltransferases", "DHHC-type acyltransferases"
Possibly the most intriguing is the reversible character of S-palmitoylation. Enzymes catalyzing palmitoylation and depalmitoylation of proteins have been characterized (58, 59).
Palmitate is transferred onto the thiol group of cysteine from
cytosolic palmitoyl-CoA by palmitoyl acyltransferases, enzymes
containing the zinc finger DHHC domain named after the highly conserved
Asp–His–His–Cys peptide (Figure 1).
This is a two-step reaction comprising transient autoacylation of zDHHC
enzymes and transfer of the fatty acyl chain from this intermediate to a
protein substrate (60).
In mammals, the zDHHC enzyme family consists of 24 proteins, and zDHHC
proteins are also found in other eukaryotes but not in bacteria nor are
they encoded by viral genomes. Mammalian zDHHC enzymes, each having at
least four transmembrane helices, are located in the plasma membrane,
endoplasmic reticulum, and Golgi apparatus (58).
They display some specificity toward their protein substrates and also
selectivity toward fatty acyl moieties other than palmitate, which
contributes to the heterogeneity of lipids attached to proteins, such as
viral glycoproteins described below (61).
In the opposite process, the thioester bond is cleaved by acyl-protein
thioesterases (APTs) (APT1 and APT2) and palmitoyl protein thioesterases
(PPTs) (PPT1 and PPT2), which are localized in the cytosol and in
lysosomes, respectively. APT1 and APT2 likely govern the dynamic
functional changes of S-acylation of proteins (62) while PPT1 and PPT2 depalmitoylate proteins during their degradation (63, 64).
Recently, serine hydrolases of the ABHD17 family have also been
identified as depalmitoylating enzymes, and their specific substrate
proteins determined (65, 66).
Of note, the zDHHCs, APT1/APT2, and ABHD17 proteins are S-palmitoylated themselves, and palmitoylation of zDHHCs and depalmitoylation of APT1/2 can occur in a cascade manner (57, 62).
The dynamic cycles of palmitoylation/depalmitoylation detected for
several peripheral membrane proteins are often synchronized with
intracellular trafficking of those proteins. They circulate between the
plasma membrane and the Golgi apparatus or endosomes, as exemplified by
N- and H-Ras, R7-regulator of G protein and APTs. In fact, it is
proposed that palmitoylation-dependent anchoring of APT1 in the plasma
membrane allows it to depalmitoylate H-Ras at this location, while
subsequent auto-depalmitoylation releases APT1 guiding it, alongside
H-Ras, for another round of palmitoylation at the Golgi apparatus (62, 67–69).
Cycles of palmitoylation/depalmitoylation are crucial for signaling by
distinct plasma membrane receptors and for their distribution (69–71).
Activation of TCR receptor or Fas receptor in T cells was found to
trigger quick and transient palmitoylation of Lck kinase of the Src
family (72, 73), but the exact meaning of the dynamic protein S-palmitoylation for processes triggered during the host–pathogen interaction awaits elucidation.
It is worth mentioning that although the zDDHC enzymes catalyze bulk protein palmitoylation in eukaryotic cells (74),
some proteins have a unique autopalmitoylation activity. These include
Bet3, a component of a multisubunit transport protein particle complex
involved in vesicular trafficking, TEA domain transcription factors, and
also bacterial Evf protein (75–78).
The palmitic acid residue is attached constitutively to a specific
cysteine residue of those proteins, remains buried inside a hydrophobic
pocked in their core thereby affecting the tertiary structure and, thus,
interactions with other proteins (75, 77). An exhaustive discussion on the physiology of S-palmitoylated proteins in eukaryotic cells can be found in several recent reviews (46, 79, 80).
S-Palmitoylation Is a Special Case of S-Acylation of Proteins
It has been established that, in addition to palmitate,
various other fatty acyl moieties, such as saturated stearate (C18:0) or
monounsaturated palmitoleate (C16:1), and oleate (C18:1) can be
attached via the thioester linkage to proteins. The early reports
on the heterogeneity of the fatty acyl moieties attached to cysteines
obtained by analysis of selected immunoprecipitated proteins (15, 16, 81–83)
have recently been complemented by a comprehensive proteomic analysis
of fatty-acylated proteins of macrophage-like RAW264 cells (14).
The latter study showed that an enrichment of culture medium of cells
with monounsaturated fatty acids leads to their incorporation into a
similar set of proteins as those normally modified with palmitate. Among
them, several proteins relevant to innate immune responses were found.
All these data justify the use of a broader term S-acylation rather than S-palmitoylation (Table 1). The physiological consequences of S-acylation
of proteins with individual fatty acids are slowly being revealed.
Modification of Fyn kinase with polyunsaturated fatty acid residue, such
as arachidonate (C20:4), disturbed its raft localization and, thereby,
TCR signaling (15). A heterogeneity of S-acylation
was also found in viral spike proteins, such as hemagglutinin (HA) of
influenza A virus, and E1 and E2 of Semliki Forest virus, which are
modified in host eukaryotic cells by attachment of both palmitate and
stearate (9).
In HA, stearate is attached at the transmembrane cysteine while
palmitate is attached to two cysteine residues in a membrane-proximal
region of the protein. The stearoyl chain seems to accommodate into a
groove formed by amino acids of the transmembrane helix shaping the
domain in a way that facilitates its fitting into rafts (84). S-stearoylation
of human transferrin receptor 1 at the juxtamembrane cysteine
residues(s) is a key factor of the signaling cascade controlling
mitochondrial morphology and functioning (13).
Of interest, the latter study also showed that dietary supplementation
of stearic acid reversed the deleterious effects of a genetically
determined mitochondria dysfunction in Drosophila. Taking into account that unsaturated fatty acids affect the profile of S-acylation of proteins in vitro (14, 15), it is of outmost interest whether a similar effect of unsaturated and saturated (palmitic) fatty acids could be achieved in vivo with respect to proteins of immune cells.
N- and O-Acylation of Proteins
Beside S-acylation, less frequently palmitate can
also be attached to the amine group of various amino acids (glycine,
cysteine, and lysine) giving N-palmitoylation or to the hydroxyl group of serine or threonine in a process called O-palmitoylation (Table 1). As during S-palmitoylation, also other fatty acids can be utilized in these processes named then N- and O-acylation. Thus, a type of protein N-acylation is N-myristoylation,
a frequent modification contributing to membrane anchoring of
peripheral proteins. The saturated myristate (C14:0) is transferred to
the protein from myristoyl-CoA by N-myristoyl transferase (two
isozymes in mammals). In a vast majority of cases, myristate is attached
co-translationally to the N-terminal glycine residue (after removal of
the initiator methionine) via an amide linkage (Table 1). Like most lipidations, this modification is irreversible. Several viral proteins are N-myristoylated,
such as Gag of HIV-1 crucial for budding of newly formed virions from
plasma membrane rafts of host cells, and proteins of parasitic protozoa Plasmodium falciparum, Trypanosoma brucei, and Leishmania donovani (causing malaria, African sleeping sickness, and leishmaniosis, respectively). For this reason, N-myristoyl transferase is considered a potential drug target in the therapy of these diseases (17–19, 85, 86)
(My Comments: Coronavirus does not use myristOylations! It needs not budding far away! It destroyes the host)
Data on the N- and O-palmitoylation of
proteins involved in the host–pathogen interactions are limited, but
interesting conclusions can be drawn from the information concerning
proteins taking part in other processes. N-palmitoylation of the N-terminal glycine of the α-subunit of a heterotrimeric G protein (Gαs) has been described (20) (Table 1) besides the well-known S-palmitoylation of this pivotal signaling protein. The N-palmitoylation
of Gαs is irreversible, and the enzyme responsible for this
modification is unknown. It has been speculated that S- to N-palmitoyl migration can occur both in vivo and also in vitro during mass spectrometry analysis (20, 87).
This suggests that caution is needed in interpreting results of this
methodological approach, which is used with increasing frequency to
study fatty acylation of proteins in immune cells (see next sections).
Probably the best-characterized is the N-palmitoylation of the
N-terminal cysteine residue of hedgehog proteins (sonic, Indian, and
desert in mammals). It determines secretion of these proteins, which
regulate embryonic patterning (Table 1). Secreted Wnt and ghrelin proteins are examples of O-acylation of serine residues with unusual fatty acid residues such as palmitoleate (C16:1) and octaonoate (C8:0) (Table 1). The fatty acylation of hedgehog, Wnt, and ghrelin is catalyzed by enzymes from the multipass membrane-bound O-acyl transferases family (31).
Besides these unusual fatty acid residues, attachment of palmitate to
serine and threonine residues is found in secreted venom toxins of the
spider Plectreurys tristis, which selectively target neuronal ion channels (88). Also histone H4 is O-palmitoylated at a serine residue in the nucleus by acyl-CoA:lysophosphatidylcholine acyltransferase (25) (Table 1). The latter is of special interest in the context of innate immune responses since histone H4 O-palmitoylation
regulates transcriptional activity, which is the final outcome of the
pro-inflammatory signaling pathways triggered by receptors of the innate
immune system.
Special attention should be devoted to ε-N-acylation consisting in the attachment of a fatty acid residue to the side chain of lysine by amide linkage (Table 1). ε-N-myristoylated
are interleukin 1α (IL-1α) and tumor necrosis factor α (TNFα), the
pro-inflammatory cytokines crucial in combating bacterial infections (22). The enzyme(s) catalyzing this reaction is unknown, but it has been established that sirtuins reverse this modification (89). The ε-N-acylation affects the release of TNFα by immune cells (90, 91).
Surprisingly, this rare modification is also found in toxins of
so-called RTX (repeats-in-toxin) class released by some pathogenic
Gram-negative bacteria (23, 24). We describe these cases in more detail in the following sections.
S-Prenylation, Another Common Lipidation of Proteins
Besides S-palmitoylation and N-myristoylation, S-prenylation
is another common lipidation that endows proteins with a hydrophobic
moiety and contributes to their association with membranes. This
modification relies on the posttranslational and irreversible attachment
of either farnesyl or geranylgeranyl chains to a cysteine residue in
the C-terminal CAAX box (alternatively also CC and CXC motifs) via
a thioether linkage. The process is catalyzed by protein prenyl
transferases that use polyprenylpyrophosphate as the donor of the
isoprenoid group (Table 1). In peripheral membrane proteins, the S-palmitoylation site is often located in proximity of N-myristoylation or S-prenylation
sites or a polybasic motif, which all are likely to mediate initial
weak binding of a protein to a membrane and thereby facilitate
subsequent attachment of palmitate to the protein by the integral
membrane zDHHC enzymes (31). In contrast to S-palmitoylation, data on the role of S-prenylation of proteins key to the host–pathogen interactions are scarce (92). However, since S-prenylation
is typical for the ubiquitous small GTPases of Ras superfamily, it is
vital for proper functioning of B and T cells (93, 94).
A glance at Table 1 indicates that palmitate can be covalently bound via oxyester, amide, and thioester linkages to respective amino acid residues creating an array of possible modifications. O- and N-palmitoylation of proteins seems to be stable, resembling in this regard the other common protein lipidations, N-myristoylation and S-prenylation. By contrast, there exist enzymes cleaving the thioester bond formed during S-palmitoylation. For a long time, our understanding of protein S-palmitoylation
and its dynamics was poor in comparison with other reversible protein
modifications due to technical difficulties. Only recently have these
difficulties been overcome with the introduction of methods allowing
high-throughput identification of palmitoylated proteins, also those
involved in the immune response to microbial pathogens, as discussed in
the next sections.
Methodological Progress Facilitates Detection of Protein Palmitoylation
One of the basic problems hindering studies on protein S-palmitoylation
lies in the fact that there is no identifiable consensus sequence for
the palmitoylation site that could facilitate its prediction. From the
technical point of view, the progress in a comprehensive survey of
protein S-palmitoylation was also hampered by a lack of
antibodies detecting this modification, with the sole exception of an
antibody specific to palmitoylated PSD-95 (95). A classical method used to demonstrate protein palmitoylation is based on metabolic labeling of living cells with [3H]-palmitic
acid, subsequent immunoprecipitation of a selected protein and
detection of the incorporated tritiated fatty acid by autoradiography (96).
A major disadvantage of this method is its low sensitivity. Only a
minute fraction of the radioactive palmitate is bound to proteins, the
majority being incorporated into lipids, which requires lengthy film
exposure (counting in days).
A methodological breakthrough in the identification of
palmitoylated proteins came with the development of two non-radioactive
methods based on so-called click chemistry (97–99) and acyl-biotin exchange (ABE) (74, 100).
These techniques have paved the way for high-throughput mass
spectrometry-based proteomic analysis of protein palmitoylation in
various cells and tissues and facilitated identification of new
palmitoylated proteins of both pathogens and host cells involved in the
innate immune responses.
The Click Chemistry-Based Method of Analysis of Protein Fatty Acylation
In the click chemistry-based method, cells are
metabolically labeled with a palmitic acid analog bearing an alkyne
group at the ω carbon of the fatty acyl chain, such as 17-octadecynoic
acid (17ODYA) or alk-16 (Figure 2A), and this step resembles the classic labeling of cells with [3H]-palmitic acid. However, in the click chemistry-based assay, the labeling and lysis of cells is followed by in vitro
coupling of the function group of the palmitic acid analog to a
reporter tag, which greatly enhances the sensitivity of detection of
labeled proteins (98, 99). Thus, after cell lysis, the labeled proteins are subjected to Cu(I)-catalyzed
cycloaddition known as “click” reaction with an azide-bearing detection
tag. In this step, a triazol is formed between the alkyne group in the
palmitic acid analog and the azide of the tag (Figure 2A).
The azide-bearing tags can be either fluorescent, such as
tetramethylrhodamine or dyes with infrared fluorescence, or carry a
biotin moiety. Depending on the tag used, subsequent SDS-PAGE separation
of proteins allows global visualization of palmitoylated proteins by
simple in-gel fluorescence or by blotting with a streptavidin-conjugated
reporter (98, 101, 102). Notably, proteins biotinylated via
the click reaction can also be enriched on streptavidin-coated beads
and then subjected to on-bead tryptic digestion (or in-gel digestion if
eluted from the beads) followed by identification by mass spectrometry.
Such comprehensive click chemistry-based proteomic analysis has brought
about identification of an array of palmitoylated proteins in dendritic
cells (10, 103), macrophage-like RAW264 cells (14), and T cells (99, 104, 105). Some of the S-palmitoylated
proteins newly identified in those studies, such as IFITM3 and TLR2,
are involved in the host–pathogen interactions regulating innate immune
responses (10, 103), while many others are known to contribute to adaptive immunity (99, 105), as described below. Recently, global profiling of Toxoplasma gondii
(the causative agent of toxoplasmosis) has been performed revealing
that many components of the parasite’s motility complex are
palmitoylated (106). Similar studies on Cryptococcus neoformans
(the fungus causing cryptococcal meningitis) have revealed a
contribution of specific zDHHC palmitoyl acyltransferase, called Pfa4,
to its virulence (107).
Moreover, application of analogs of various saturated and unsaturated
fatty acids confirmed the heterogeneous nature of the fatty acylation of
proteins in RAW264 cells and suggested that dietary unsaturated fatty
acids, after incorporation to proteins, can change their properties and
thereby affect the functioning of immune cells (14).
Figure 2. Detection of S-palmitoylated proteins using click chemistry and acyl-biotin exchange (ABE). (A)
Click chemistry-based method. Cells are metabolically labeled with an
alkyne-functionalized palmitic acid analog, such as 17-octadecynoic acid
(17ODYA), and after cell lysis, the click reaction is conducted with
azido-tagged biotin or fluorescent probes allowing enrichment and
detection of labeled proteins in various ways. Biotinylated proteins can
be bound on a streptavidin resin and then released using, e.g., high
concentrations of urea and SDS (108).
When a cleavable derivative of biotin, azido-azo-biotin, is used the
labeled proteins are eluted from streptavidin beads with sodium
dithionite, which cleaves the diazobenzene moiety in the linker arm of
azido-azo-biotin, and analyzed by mass spectrometry or immunoblotting (109). (B)
ABE method. Cells or tissues are lysed, free thiol groups of proteins
are blocked by alkylation, and palmitoyl moieties are released with
hydroxylamine. The newly exposed protein thiol groups are subjected to
labeling with biotin-HPDP allowing selective binding, elution, and
analysis of the originally S-palmitoylated proteins. The proteins
can also be captured without biotinylation through a direct interaction
of their thiol residues with a thiol-reactive resin (acyl-RAC
technique).
The major advantage of the click chemistry-based method is that it can reveal the time course of protein S-palmitoylation.
By using click chemistry-based labeling in the pulse-chase mode, one
can follow the dynamics of protein palmitoylation. With such an
approach, it was found that the palmitate turnover on Lck, an Src-family
tyrosine kinase, is accelerated by T cell activation (72).
Additional introduction of stable isotope labeling by amino acids in
cells (SILAC) has provided quantitative proteomic data on the dynamics
of protein palmitoylation in the cell (104, 110).
This approach revealed, rather unexpectedly, that in unstimulated T
cell hybridoma, the palmitoylation of most protein species does not
undergo turnover (104).
Another advantage of the click chemistry-based assay is its high
specificity, because the alkyne group introduced in the analog of
palmitic acid is not normally found in cells (98, 102).
The click chemistry-based methods can also be used to follow the
cellular localization of palmitoylated proteins by immunofluorescence
when combined with the proximity ligation technique (111, 112). Palmitoylation of individual proteins can also be studied after their immunoprecipitation (11, 72, 73, 98).
Despite its unquestionable success, the click
chemistry-based methods have limitations. They will detect only those
proteins that undergo palmitoylation during the period of the metabolic
labeling of cells. One should also bear in mind that the palmitic acid
analog can be incorporated at S-, N-, and O-palmitoylation sites alike (111, 112).
In addition, although 17ODYA (alk-16) is preferentially used to mimic
palmitoylation of proteins, it can also be incorporated with low
efficiency at N-myristoylation sites of proteins (98, 99). Another group of proteins that will be labeled with the palmitic acid analog but are not S-palmitoylated are those bearing the glycosylphosphatidylinositol (GPI) anchor (85, 113).
Most of these limitations can be overcome using various fatty acid
reporters, inhibitors, and by exploiting the sensitivity of the
thioester bond to hydroxylamine treatment. Given the large variety of
chemical reporters preferentially mimicking distinct fatty acids, recent
years have witnessed a plethora of chemistry-based proteomic studies
not only on palmitoylated but also myristoylated proteins and proteins
bearing the GPI anchor, including those of pathogens and immune cells (10, 14, 85, 86, 114).
The ABE Method Reveals Protein S-Acylation
The ABE method can be used as a complement to the click
chemistry-based approach in cell studies but unlike the latter it is
uniquely suitable for studying whole tissues. ABE does not require
metabolic labeling of proteins in living cells, thus some of the
abovementioned limitations and difficulties do not apply. The ABE method
relies on in vitro exchange of thioester-linked palmitate to a
derivative of biotin which allows subsequent affinity purification of
the resulting biotin-labeled proteins on streptavidin-coated beads
(Figure 2B).
The first step of the ABE involves lysis of cells or tissues followed
by irreversible blockage of free thiol groups in the solubilized
proteins by alkylation, most often with N-ethylmaleimide. Subsequently, the thioester bonds existing in S-palmitoylated
proteins are broken with hydroxylamine, releasing palmitoyl moieties.
The newly exposed thiol groups can now be tagged with
sulfhydryl-reactive derivatives, such as biotin-HPDP, forming disulfide
bonds with thiols. The biotinylated proteins are subsequently captured
on streptavidin-coated beads and eluted with agents that reduce the
disulfide bond between the protein and biotin-HPDP, such as
β-mercapthoethanol, DTT, or TCEP (49, 74, 115, 116).
As an alternative to biotinylation, in the so-called acyl-RAC
technique, the newly exposed protein thiol groups in
hydroxylamine-treated cell lysates are captured on a resin containing
sulfhydryl-reactive groups (117).
In both ABE and acyl-RAC, the eluted proteins can be separated by
SDS-PAGE and visualized by gel staining or immunoblotting, or identified
by mass spectrometry. Furthermore, when the hydroxylamine-released
palmitoyl moieties are exchanged for a polyethylene glycol-maleimide
derivative of a distinct molecular weight, a shift in-gel migration of
tagged proteins is observed reflecting the number of fatty acyl residues
originally S-bound to the protein (118, 119).
The ABE method has so far been used successfully for proteomic profiling of S-acylated proteins in immune cells, such as RAW264 cells (48), several types of blood cells, such as platelets, primary T cells, and immortalized B cells (120–122), pathogenic microorganisms such as T. brucei and T. gondii (123, 124), and tissues (125, 126).
To quantify the aberrations in protein palmitoylation in a mouse model
of Huntington’s disease, whole animal stable isotope labeling of mammals
(SILAM) was applied followed by tissue isolation and ABE procedure (127).
In another approach, for quantitative analysis of the T-cell
palmitoylome, ABE was combined with labeling of proteins with various
oxygen isotopes during their digestion with trypsin before mass
spectrometry analysis (122).
In addition, preselection of tryptic peptides obtained by ABE on
streptavidin-coated or sulfhydryl-reactive resins greatly facilitates
the identification of S-acylation sites by mass spectrometry (49, 110, 117).
Some aspects of the ABE method deserve a comment. Since
the assay relies on the sensitivity of thioester bonds to hydroxylamine,
ABE detects all S-acylation without distinguishing between S-palmitoylation
and the other cases. Furthermore, there is a possibility of
false-positive detection of proteins bearing a thioester linkage with
compounds other than fatty acyl residues, such as ubiquitin in the E2
ubiquitin conjugase Ubc1 (115).
Another source of false-positives is proteins in which free thiol
groups were not completely alkylated before biotinylation. On the other
hand, insufficient deacylation of bonafide fatty-acylated proteins with hydroxylamine results in their absence in the final sample (116).
In summary, the click chemistry-based method relies on
metabolic labeling of cells with a palmitic acid analog which
incorporates into proteins and next tagging it with reporter molecules
greatly enhancing the sensitivity of detection. It only reveals proteins
undergoing S-palmitoylation during metabolic labeling of cells
and allows revealing turnover of this modification. By contrast, the ABE
method is based on direct binding of sulfhydryl-reactive derivatives to
thiol groups of cysteines unraveled by hydroxylamine treatment after
lysis of cells or tissues. It allows the investigation of the whole but
static palmitoylome. A comparative proteomic study of protein
palmitoylation in P. falciparum found that the sets of proteins identified using these two approaches overlapped in 57.2% (113),
indicating that they provide complementary data on the cellular
palmitoyl proteome. Thanks to the application of the click chemistry-
and ABE-based methods numerous new palmitoylated proteins have been
identified. In 2015, a SwissPalm database was launched, (128)
which provides an excellent, manually curated resource of information
on palmitoylated proteins, palmitoylation sites, etc., available at http://swisspalm.epfl.ch/.
All these efforts have greatly furthered our knowledge on molecular
mechanisms regulating diverse aspects of cell functioning, including
host–pathogen interactions and progress of infectious diseases, as
highlighted below.
Palmitate as a Component of Proteins and Lipids Related to Bacterial Pathogenicity
Bacteria lack protein palmitoyl acyltransferases of the zDHHC family and, therefore, are essentially devoid of S-palmitoylated
proteins. Yet, they have developed unique mechanisms utilizing fatty
acids, such as palmitic acid, to modify their glycolipids and proteins.
These modifications augment infectivity and help bacteria evade
recognition by the host innate immune system. For example, the vast
majority of Gram-negative bacteria produce lipopolysaccharide (LPS) as a
part of their outer membrane. LPS is composed of the variable
polysaccharide O-antigen and more-conserved lipid A containing
two glucosamine residues hexa-acylated with hydroxymyristic, myristic,
and lauric acid. Lipid A is recognized by CD14 protein and TLR4 receptor
complexed with MD2 protein on the plasma membrane of the host immune
and some non-immune cells. Activation of TLR4 triggers strong
pro-inflammatory reactions aiming at eradication of the bacteria, but
when exaggerated, eventually leading to sepsis (129).
Incorporation of an additional palmitoyl chain into lipid A markedly
diminishes its ability to activate TLR4 and to induce the host
pro-inflammatory responses, which is correlated with an increased
survival of bacteria forming a biofilm (130, 131). This strategy is utilized among others by Salmonella typhimurium, a causative agent of gastroenteritis, by Bordetella bronchiseptica, a respiratory pathogen of human and other mammals, and by Yersinia pestis causing plague (132, 133).
The formation of the extra-acylated LPS relies on the transfer of
palmitate from phospholipids onto the hydroxymyristate chain at position
2 of glucosamine of lipid A. The reaction is catalyzed by lipid A
palmitoyltransferases (PagP in Salmonella and its homologs in other bacteria) localized in the outer membrane of these pathogens (134, 135).
In addition to causing steric hindrance preventing the binding to the
TLR4/MD2 complex, the hepta-acylation of LPS also protects bacteria from
the lytic activity of cationic antimicrobial peptides, most likely by
reducing the fluidity of the bacterial outer membrane (136, 137).
Apart from being incorporated into LPS in diverse
bacteria, palmitate has also been found to modify a virulence factor of
Gram-negative Erwinia carotovora, the Evf protein. The palmitoyl chain is linked via a thioester bond to the Cys209 residue at the center of Evf, plausibly by a self-palmitoylating activity of the protein. E. carotovora is a phytopatogen using insects such as Drosophila as vectors for dissemination between plants. The palmitoylation of Evf is required for infectivity of E. carotovora
and its persistence in the insect gut, however, its mode of action of
unknown. It has been speculated to be linked with an ability of Evf to
associate with lipid bilayers, but the lack of similarities between Evf
and any other bacterial protein of known function makes prediction on
this subject difficult (79).
A number of bacterial toxins of so-called RTX class
released during infection of mammals by pathogenic Gram-negative
bacteria undergo ε-N-acylation of the side chain of internal lysines. These toxins include adenylate cyclase of Bordetella pertussis, acylated with palmitic acid, and α-hemolysin of extraintestinal (uropathogenic) Escherichia coli,
acylated with myristic acid and also 15- and 17-carbon fatty acids. The
acylation is catalyzed by an endogenous bacterial acyltransferase
which, unlike its eukaryotic counterparts, transfers the acyl chain not
from acyl-CoA but from acyl-carrier protein. The acylated toxins
secreted by the bacteria bind to the plasma membrane of the host cells,
oligomerize and form pores causing cell lysis. In the case of the toxin
of B. pertussis, essential is also the delivery of the adenylate
cyclase moiety to the cell interior. Acylation is required for virulence
possibly being involved in oligomerization of the toxins (23, 24, 138).
Although lacking S-palmitoylated proteins (with
the single known exception of Evf), bacteria express a wide range of
membrane-bound proteins modified by a complex lipidation at the
N-terminus, with palmitate frequently being a component of the lipid
moiety (139, 140).
The bacterial lipoproteins are synthesized in a multistep process
catalyzed by a unique set of lipoprotein processing enzymes, Lgt, LspA,
and Lnt, absent in eukaryotic cells. The formation of these lipoproteins
begins with the attachment of a diacylglycerol via a thioester
bond to a cysteine residue located in the so-called lipobox motif of the
signal sequence of the transmembrane lipoprotein precursor. The signal
sequence is then cleaved next to the lipid-modified cysteine leaving it
at the N-terminus of the mature protein (141). In Gram-negative and less frequently also Gram-positive bacteria, a third fatty acid residue is additionally attached via an amide linkage to the amino group of the cysteine in a reaction analogous to the N-acylation of hedgehog proteins (see Table 1).
This di- and tri-lipidation ensures membrane anchoring of the
lipoproteins. All such lipoproteins of Gram-positive bacteria are
exposed to the milieu while in Gram-negative bacteria some face the
periplasm. The lipoproteins of Gram-positive bacteria, e.g., Streptococcus pneumoniae (causing pneumonia), Mycobacterium tuberculosis (tuberculosis), and Gram-negative bacteria, such as Neisseria meningitidis (meningitis), Y. pestis (plague), the spirochaete Borrelia burgdorferi (Lyme disease) and Treponema pallidum
(syphilis) are crucial for their virulence. They control several
aspects of the host–pathogen interactions, like adhesion and entry to
host cells, protection against proteolysis and oxidative stress in the
host cell, and regulation of expression of genes encoding cytokines both
during initiation and progress of the disease (140–142).
The surface exposure of the lipoproteins allows their involvement in
the host cell invasion while on the other hand forming the so-called
pattern signal recognized by the TLR2 receptor, which triggers the
pro-inflammatory responses helping to combat the bacteria (143). Of interest, TLR2 is S-palmitoylated,
as discussed below. The involvement of lipoproteins in pathogenesis
fuels studies on their properties. One such recent work employing click
chemistry to profile the lipoproteins of E. coli identified 88 lipoproteins with high/medium confidence, 70% of them predicted before by bioinformatics analysis (144).
Notably, in that study a 14-carbon alkynyl fatty acid analog alk-14
rather than alk-16 was preferentially incorporated into the
lipoproteins, contradicting earlier studies using gas chromatography and
TLC, which found that palmitate was predominantly used for bacterial
protein modification (139).
Further studies are required to establish whether the fatty acid found
in lipoproteins varies depending on culture conditions or is species
specific. For example, 17ODYA labeling for click reaction confirmed
incorporation of palmitate into pallilysin (Tp0751), a lipoprotein of T. pallidum.
Pallilysin is a metalloprotease that degrades human fibrinogen and
laminin. It is suggested that its exposure on the bacteria surface
enables degradation of host structural proteins to facilitate rapid
dissemination of this highly invasive pathogen (140).
Bacteria occasionally high-jack the palmitoylation
machinery of host cells to modify the environment so as to favor their
internalization, survival, and replication inside the cells. Bacillus anthracis (the causative agent of anthrax) is an example of such bacteria that modify S-palmitoylation
of host proteins to their ends. The anthrax toxin produced by this
pathogen binds to the TEM8 and CMG2 (capillary morphogenesis protein-2)
proteins which, under physiological conditions, are involved in
cell–cell and cell–extracellular matrix interactions. They are S-palmitoylated at multiple (two to four) cysteines (54). The S-palmitoylation
of TEM8 was found to inhibit its association with plasma membrane rafts
preventing its ubiquitination by the raft-associated E3 ubiquitin
ligase Cbl. The binding of anthrax toxin drives association of the
receptor-toxin complexes with rafts possibly correlated with
depalmitoylation of the receptor. This allows subsequent ubiquitination
of the receptor, an uptake of the receptor/toxin complexes in a
clathrin-dependent manner and eventual delivery of the toxin to
endosomes. These events are facilitated by S-palmitoylation of partner(s) of the receptors, most likely including kinases of the Src family (54, 145, 146).
While B. anthracis utilizes palmitoylated host
proteins to induce its internalization, a growing body of data suggests
that also bacterial proteins can undergo S-palmitoylation inside
the host cells. This type of modification concerns so-called effectors,
bacterial proteins that are injected into the host cell cytoplasm either
across the plasma membrane or the membrane of vesicles enclosing
internalized pathogens, with the help of their secretion systems. These
are secretion systems type III and type IV, homologs of which have been
described for pathogens and symbionts of mammals, insects, and plants (147, 148). The bacterial effectors can be S-palmitoylated
to reach host cell membranes and thereby accumulate at a location most
suitable for their activity. Application of the click chemistry-based
method utilizing an analog of palmitic acid (alk-16) for cell labeling
has revealed S-palmitoylation of two effector proteins of Salmonella enterica, such as SspH2 and SseI (149). S. enterica
invades gut endothelial cells and is a leading cause of gastroenteritis
and typhoid fever. SspH2 carries an E3 ubiquitin ligase domain while
SseI shows sequence homology to bacterial proteins that have a deamidase
activity, and inhibits migration of Salmonella-infected cells. The latter activity requires S-palmitoylation of SseI. Both proteins are stably S-palmitoylated,
most likely by zDHH3 and zDHH7 of the host and bind to the plasma
membrane in a palmitoylation-dependent manner (149). Also two effector proteins of the IpaH family of Shigella spp. were found to be S-palmitoylated
in that study, suggesting that this modification can control the
activity of effector proteins of other pathogens as well (149). Indeed, GobX and LpdA, effector proteins of Legionella pneumophila, the causative agent of Legionnaires’ disease invading macrophages and lung endothelial cells, are S-palmitoylated
as was found recently using click chemistry. LpdA is a phospholipase
hydrolyzing various phosphatidylinositols while GobX is an E3 ubiquitin
ligase. GobX is targeted in a palmitoylation-dependent manner to the
Golgi apparatus, and LpdA to the plasma membrane and a subset of
intracellular vesicles (150, 151). Thus, the diversified subcellular localization of bacterial effector proteins reflects that of eukaryotic proteins.
It is worth noting that global profiling of acylated
proteins with the application of click chemistry and an
alkyne-functionalized analog of myristic acid, alk-14, for cell labeling
was effective in reveling the mechanism of action of Shigella flexneri
effector protein IpaJ of type III secretion system. This is a unique
protease that cleaves off the N-terminal myristoylated glycine. This
proteolytic demyristoylation activity of IpaJ is specific toward
Golgi-associated ARF/ARL family of GTPases regulating cargo transport
through the Golgi apparatus, inhibition of which is apparently pivotal
for virulence of the bacteria causing diarrhea in humans (152).
In addition to the S-palmitoylation of the effectors of pathogenic bacteria of mammals mentioned earlier, double acylation, N-myristoylation and S-palmitoylation, has been reported of the so-called avirulence (Avr) proteins (effectors of type III secretion system) of Pseudomonas syringae, a causative agent of diverse plant diseases. Among them, AvrRpm1 and ArvB are N-myristoylated and S-palmitoylated
by host acyltransferases at neighboring glycine and cysteine residues
localized at the N-terminus of the proteins (similarly to eukaryotic
kinases of the Src family), while in AvrPphB and two AvrPphB-like
effectors—ORF4 and NopT, the double acylation motif is exposed after
auto-cleavage of the proteins (similarly to some eukaryotic proteins
cleaved by caspases). The acylation of the Avr proteins ensures their
anchoring in the host plasma membrane, which is required for their
functioning. In disease-susceptible plants Avr proteins contribute to
successful infection; however, in plants expressing host resistance (R)
genes they trigger plant defense signals, in both cases engaging plasma
membrane-associated host proteins (153, 154).
The importance of palmitoylation of bacterial effector
proteins for their infectivity is only beginning to be uncovered, in no
small part owing to the development of the click chemistry-based method
for detection of this protein modification. However, the strategy of
high-jacking the host palmitoylation machinery to modify own proteins
seems to be much more commonly employed by viruses.
Protein Palmitoylation in Viral Infections
Viruses do not encode palmitoyl acyltransferases but
exploit extensively the host palmitoylation machinery to modify their
proteins essential for infection of host cells and own replication. In
fact, S-palmitoylation of proteins was discovered in 1979 as a
modification of envelope glycoproteins of Sindbis virus and VSV. In
those studies [3H]-palmitic acid was used for metabolic
labeling of virus-infected cells and labeled proteins were identified by
autoradiography (12, 155). Subsequently, a number of other viral proteins have been found to be palmitoylated using this approach.
The most-studied group of viral palmitoylated proteins
is those found in enveloped viruses, i.e., viruses covered by a lipid
bilayer obtained during their replication from a membrane of the host
cell, such as the plasma membrane or endoplasmic reticulum. Influenza
virus, HIV-1, hepatitis C virus (HCV), and herpes simplex virus (HSV)
are the best known enveloped viruses. The envelope is rich in
transmembrane, often S-palmitoylated, glycoproteins called
spikes, which can bind to cognate receptors on the host cell plasma
membrane triggering endocytosis of the virion, mediate subsequent fusion
of the viral and cellular membranes allowing entry of the viral genome
to the cytoplasm, and are also involved in the budding of newly formed
virus particles from the cell. An example of such multifunctional
palmitoylated transmembrane glycoproteins is HA present in the envelope
of influenza virus together with another palmitoylated transmembrane
protein, the matrix protein M2, which forms a proton channel earning the
protein the name viroporin. As mentioned earlier, HA of influenza A
virus is S-stearoylated and S-palmitoylated, respectively,
at one cysteine residue located in the transmembrane domain of HA and
two cysteines found in the cytoplasmic (intraviral) tail in close
proximity to the membrane (156).
On the other hand, M2 is S-palmitoylated on the amphiphilic helix located in the cytoplasmic part of the protein. Due to the S-palmitoylation and the presence of a cholesterol-binding motif the helix bends toward and associates with membranes (157, 158).
During infection, HA binds to sialic acid residues of glycans localized
on the surface of airway and alveolar epithelial cells. The bound
virions are endocytosed and next the viral and endosome membranes fuse.
The membrane fusion is driven by HA, which undergoes conformational
changes induced by low pH of endosomes. Acidification of endosomes
activates also the M2 proton channel activity, protons entering viral
core facilitate dissociation of the viral genome which then moves to the
nucleus where RNA replication occurs. The S-palmitoylation of HA
is required for the fusion of the viral and endosome membranes at least
in some subtypes of the virus while the ion channel activity of M2 is
not dependent on its S-palmitoylation (159)
.
Newly synthesized viral proteins and RNA are assembled into virions in
the plasma membrane rafts which merge into lager platforms crucial for
the virion assembly and budding off. The triple fatty acylation of HA is
required for its targeting to plasma membrane rafts (160, 161).
Besides S-palmitoylation, also the amino acid sequence of the
transmembrane domain of HA determines its association with rafts (45). On the other hand, among the amino acids of the cytoplasmic tail of HA no other than the two S-palmitoylated
cysteines are required for viral assembly and replication, although it
is still not clear whether raft targeting (in cooperation with the
transmembrane fragment) is the only mechanism of their participation. It
is proposed that they affect conformation of the HA tail controlling
its interaction with structural matrix protein M1 lying beneath the
viral envelope (162, 163). The budding off of the virion is facilitated by M2 which localizes at the edges of rafts as a result of a combination of its S-palmitoylation,
cholesterol binding, and properties of the transmembrane fragment. M2
protein can create a “wedge” altering membrane curvature thereby
facilitating membrane scission and release of the virion (157, 164).
The influenza virus S-palmitoylated proteins are the archetype for many other viral proteins. Thus, S-palmitoylated
spike glycoproteins include S-protein of coronaviruses (e.g., severe
acute respiratory syndrome virus), the fusion (F) protein of
paramyxoviruses (e.g., measles virus), Env of retroviruses [e.g., HIV-1,
feline immunodeficiency virus (FIV)], and filoviruses (e.g., Ebola).
Other viral proteins modified with palmitate are viroporins, such as E
protein of coronaviruses, and also peripheral membrane proteins or
nucleocapsid proteins absent in influenza virus. It has been found that S-palmitoylation
of F13L, a peripheral protein of the envelope of vaccinia virus,
controls the association of the protein with intracellular membranes,
thereby the formation of the envelope (165).
The core protein of the nucleocapsid of HCV resides on the surface of
lipid droplets and binds in a palmitoylation-dependent manner to
membranes of the droplet-associated endoplasmic reticulum. Subsequently,
it recruits viral proteins and newly synthesized RNA for viral particle
formation (166). Besides the interest in the role of viral protein S-palmitoylation for infectivity and possible use of host zDHHC enzymes as targets of anti-influenza drugs (167),
viral proteins often serve as a model to study the consequences of
fatty acylation for protein functioning and localization in distinct
membrane domains (see S-Palmitoylation of Proteins and Its
Influence on Protein Localization, Trafficking, and Stability of this
review). Readers are referred to recent exhaustive reviews that consider
these topics (36, 84, 168)
while we will focus here on the recent advances in the field of viral
protein palmitoylation brought about mainly by proteomic studies.
The click chemistry-based approach has led to the identification of S-palmitoylation
in the cytoplasmic domain of the transmembrane spike protein Env of
FIV, considered to be the cat equivalent of HIV-1. Env comprises three
transmembrane gp41 glycoproteins and three associated gp120 which bind
to CD4 receptor and coreceptors on the surface of T lymphocytes allowing
fusion of the viral envelope and the plasma membrane and entry of viral
capsid. Four cysteines in FIV Env are S-palmitoylated vis-a-vis
two found in the Env of HIV-1. The two most membrane-proximal
cysteines, 804 and 811, are required for the FIV membrane-fusion
activity and incorporation of Env into virions (169), in agreement with the importance of Env S-palmitoylation for virion assembly of some HIV-1 strains (170–172). The assembly of HIV-1 virions takes place in plasma membrane rafts and is driven by N-myristoylated
Gag protein which anchors and oligomerizes preferentially in these
plasma membrane domains due to the presence of the fatty acyl chain (18).
The development of click chemistry-based methods allowed
for the first time global profiling of acylated proteins in
virus-infected cells. In addition to identifying acylated viral proteins
this approach has also revealed how the viral infection modulates the
acylation pattern of the host cell proteins. Thus far, click chemistry
has been used to study protein myristoylation and palmitoylation in
cells infected with HIV-1 and with HSV. In the latter case, the standard
metabolic labeling with alkyne-functionalized myristic and palmitic
acid analogs followed by click chemistry and mass spectrometry was
combined with SILAC to discern between the changes in the extent of
protein acylation and those in their abundance following viral
infection. This approach allowed an elaborate quantitative analysis of
host protein acylation and has revealed an overall downregulation of the
level of both host protein modifications in infected cells. While the
decreased content of myristoylated proteins resulted mainly from
suppression of host protein synthesis, the drop in several S-palmitoylated
proteins ensued from the inhibition of their palmitoylation in infected
cells. The affected proteins were localized mainly to the plasma
membrane and the Golgi apparatus and were involved in vesicle-mediated
transport and ion transport. In addition, the study has expanded the
list of HSV-encoded acylated (mostly palmitoylated) proteins that play
different functions in the viral cycle, such as gE, gI, gK, US2, and US3
(110).
Similar results pointing to global changes of host protein acylation
were obtained upon analysis of protein myristoylation and palmitoylation
in cells infected with HIV-1. In that study, the cells were labeled
with analogs of palmitic or myristic acid tagged with an azide moiety
for click chemistry reaction; however, the following mass spectrometry
analysis did not address the relation between changes of protein
acylation vs. alteration of protein level. The study identified 17
palmitoylated and 7 myristoylated proteins significantly differing in
abundance between HIV-1 infected and uninfected cells. Several of the
proteins affected by the infection were of host origin. The abundance of
myristoylated proteins was in general increased while that of the
palmitoylated ones—decreased in infected cells (173).
In other words, the two studies have revealed that HSV and HIV-1 not
only encode proteins that are acylated in the host cell but also alter
the palmitoylation of host proteins, likely to adapt the cellular
environment to favor their replication and budding. The majority of the
acylated proteins affected by HIV-1 or HSV infection had not been
described earlier in this context; therefore, further studies on these
proteins could be crucial for better understanding of viral infection.
Thus, the click chemistry-based approach has been highly effective in
revealing changes of the host protein palmitoylation and opening new
possibilities for the identification of novel antiviral drug targets.
Palmitoylation of Host Proteins Involved in Antibacterial and Antiviral Defense
The innate immune responses are the first line of active
defense against microbial infections. The application of click
chemistry-based and ABE methods and their use for large-scale analysis
of protein palmitoylation in murine dendritic CD2.4 cells (10, 103), and murine macrophage-like RAW264 cells (14, 48) complemented by proteomic analysis of the raft fraction of those cells (47)
have contributed significantly to the understanding of the role of
palmitoylation of host receptors and signaling proteins involved in
innate immune responses. Thus, the palmitoyl proteome analysis of murine
dendritic cells unraveled S-palmitoylation of TLR2, a receptor
expressed in cells of myeloidal lineage, which heterodimerizes with TLR1
or TLR6 to bind bacterial tri- or diacylated lipoproteins,
respectively, and also other microbial components, such as glycolipids
(e.g., lipoarabinomannan) of Mycobacterium and yeast zymosan (174).
Besides TLR2, two other human TLRs out of 10 ectopically expressed in
HEK293 cell, flagellin receptor TLR5, and TLR10, a unique TLR negatively
regulating the pro-inflammatory activity of TLR2, were also found to be
palmitoylated. The S-palmitoylation site of human TLR2 was
mapped to Cys609 adjacent to its transmembrane domain. The modification
was present in unstimulated cells and was linked with up-regulation of
the cell surface localization of TLR2. Mutation of Cys609 abolished the
ability of the receptor to induce pro-inflammatory signaling in response
to microbial ligands of TLR2 (10). Further studies are needed to reveal whether S-palmitoylation of TLR2 controls its association with rafts as sites of TLR2 activation (175) and/or affects endocytosis of the receptor, as found for the anthrax toxin receptor (54).
One of the most extensively studied TLRs, TLR4 activated
by bacterial LPS, is not palmitoylated. Yet, saturated fatty acids have
been indicated to trigger pro-inflammatory signaling of TLR4. Thus, the
TLR4/MD2 receptor complex is involved in the pro-inflammatory outcome
of a diet rich in palmitic acid, as was found when analyzing markers of
inflammation in the heart and adipose tissue of high fat diet-fed mice (176, 177).
The molecular mechanisms underlying the pro-inflammatory properties of
palmitic acid can involve its influence on the plasma membrane lipid
order, hence raft organization, in a way that facilitates translocation
of TLR4 (and TLR2) toward rafts (178, 179). Palmitic acid also directly binds to the TLR4-associated MD2 protein (177, 180).
An influence of palmitic acid on sphingomyelin/ceramide metabolism,
which enhances the LPS-induced responses, has also been considered (181).
Recent proteomic studies based on 17ODYA labeling of RAW264
macrophage-like cells followed by click chemistry have revealed that
stimulation of cells with LPS induces profound changes of the abundance
of palmitoylated proteins (182). The data are in agreement with earlier findings showing that LPS induces accumulation of S-palmitoylated Lyn kinase in the raft-enriched fraction of cells, allowing it to downregulate TLR4 signaling (11). One of the upregulated S-palmitoylated
proteins was type II phosphatidylinositol 4-kinase IIβ, which
phosphorylates phosphatidylinositol to phosphatidylinositol
4-monophosphate (PI4P) (Comment: nuclear direction?). It was shown that palmitoylation determines the
involvement of the kinase in LPS-induced signaling (182). These data suggest that S-palmitoylated
proteins, including enzymes catalyzing phosphatidylinositol synthesis
and turnover, are important factors affecting the pro-inflammatory
responses triggered by LPS.
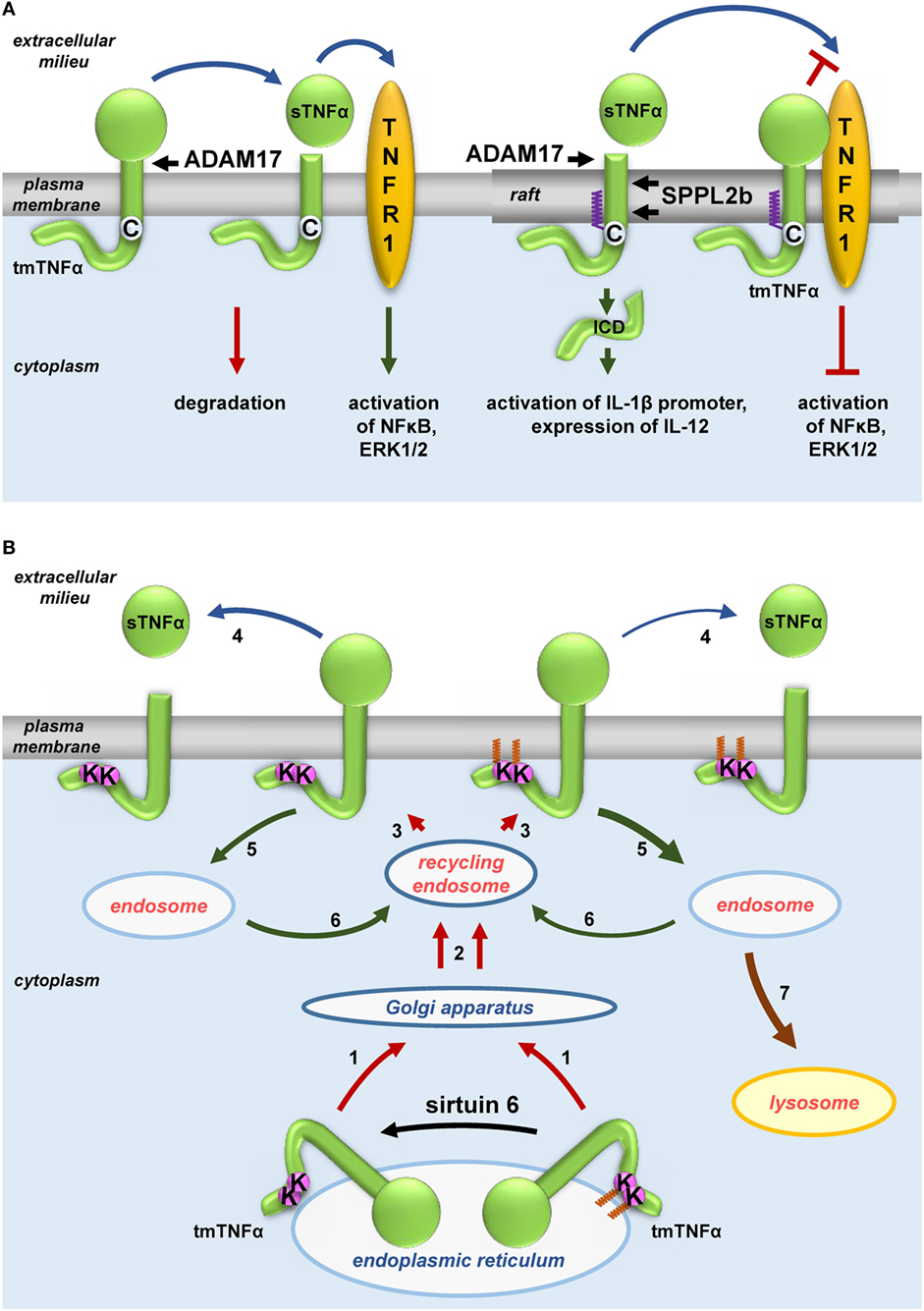
Notably LPS induces production of TNFα, a pro-inflammatory cytokine that is S-palmitoylated
itself. TNFα is synthesized as a transmembrane 27-kDa precursor
(tmTNFα) transported from the endoplasmic reticulum to the plasma
membrane through the Golgi apparatus and recycling endosomes (183). Human tmTNFα is S-palmitoylated
at Cys30 located at the boundary between its transmembrane and
cytosolic fragments, as was found independently by radiolabeling and by
labeling with 17ODYA followed by click chemistry (184, 185). Poggi et al. (185) arrived at a complex model explaining how the S-palmitoylation of TNFα affects its activity (Figure 3A).
The modification was shown to favor the association of tmTNFα with
rafts. Upon cell activation, the extracellular domain of tmTNF is
cleaved by ADAM17 metalloproteinase whereupon the soluble TNFα (sTNFα)
is released to the extracellular milieu and activates TNF receptor
(TNFR) 1 and TNFR2. As ADAM17 localizes to both non-raft and raft
regions of the plasma membrane, the S-palmitoylation of tmTNFα does not affect its cleavage and production of the soluble cytokine. However, S-palmitoylated
tmTNFα interacts with TNFR1 in rafts thereby reducing the binding of
sTNFα and consequently reducing the sensitivity of the cell to this
cytokine. In addition, the fragment of tmTNFα which remains after the
release of sTNFα in rafts if further processed by intramembrane SPPL2a
and 2b proteases giving rise to ICD (intracellular domain) of an own
biological activity. By contrast, the non-raft fragment of the
ADAM17-cleaved tmTNFα is rapidly degraded (185).
The transport and maturation of TNFα are also regulated by another posttranslational acylation, ε-N-myristoylation (22). As shown in Figure 3B,
myristic acid residues are attached to two lysines (Lys19 and 20) of
human tmTNFα. This modification is reversed by sirtuin 6 catalyzing the
demyristoylation. Depletion of sirtuin 6 decreases the release of sTNFα
since the ε-N-acylated TNFα precursor is redirected to and accumulates in lysosomes (90, 91). It is worth noting that exogenous palmitic acid stimulates the ε-N-myristoylation of tmTNFα, thereby reducing the release of sTNFα in favor of accumulation of tmTNFα in lysosomes (90, 91).
This somehow surprising anti-inflammatory effect of palmitic acid can
be explained by competitive binding between long-chain fatty acids (in
this case, palmitic) and myristoylated substrates of sirtuin 6 found in vitro—(89) and adds a new dimension to the potential effects of palmitic acid.
S-palmitoylation of host proteins is also vital
in antiviral defense. Viral nucleic acids, which are recognized by
several TLRs and also cytoplasmic pattern-recognition receptors, induce
robust production of type I interferons (IFNs), mainly INFα and IFNβ.
The IFNα and IFNβ released from cells which first encounter viruses,
e.g., dendritic cells, induce an antiviral reaction in an autocrine and
paracrine manner upon binding to plasma membrane IFNα/β receptor (IFNAR)
consisting of subunits 1 and 2. Both human IFNAR subunits are S-palmitoylated, as has been found by classical radiolabeling. The S-palmitoylation
of IFNAR1 on Cys463, localized near the cytoplasmic end of the
transmembrane domain, is required for downstream activation of STAT1 and
STAT2 and the following transcription of IFNα-activated genes (186).
Among the IFN-induced proteins, some have been shown to be
palmitoylated, using click chemistry and ABE. They include the
immunity-related GTPase Irgm1, BST2 also known as tetherin, and IFITM1
and 3 (10, 104).
IFITMs are potent restriction factors against a wide range of enveloped
viruses, e.g., influenza, West Nile, dengue, and Zika viruses (187, 188).
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4295558/
- Filoviruses and the Severe Acute Respiratory Syndrome CoronavirusThe IFITM proteins also efficiently restrict the filoviruses EBOV and Marburg virus (MARV) as well as severe acute respiratory syndrome coronavirus (SARS-CoV), as demonstrated by overexpression and short hairpin RNA (shRNA)-depletion studies using both infectious viruses and pseudoviruses (21). Although these viruses are from different families, they share a dependency on the enzymatic activities of lysosomal cathepsins to activate their fusion proteins (42, 43). Thus, they are thought to fuse later in the endocytic pathway than IAV or flaviviruses. IFITM expression does not detectably change the level of cathepsin L activity in cell lysates or the surface expression of the SARS-CoV receptor ACE2 (21, 44). SARS-CoV and filoviruses also share a pattern of restriction that is different from that of IAV. For example, SARS-CoV and filoviruses appear to be more sensitive to IFITM1 than is IAV, and unlike IAV, they can be efficiently restricted by murine Ifitm6 expressed in human cells (21). These studies provide the first suggestion that different IFITM proteins might specialize in targeting different viruses.
IFITMs localize primarily to endolysosomal membranes where they inhibit
viral replication by blocking their fusion with these membranes and
also facilitate virus degradation (187).
The exact mechanism of this antiviral activity is not clear, but it
seems to rely on a perturbation of the organization of endolysosomal
membranes. This can be linked with the intramembrane topology of IFITMs
and their S-palmitoylation. IFITM1 and 3 likely possess two loops
embedded in but not spanning the membrane with both the N- and
C-termini facing the cytoplasm (55, 189). S-palmitoylation
of conserved cysteine residues adjacent to these loops, Cys71, 72, and
105 in murine IFITM3, contributes to the membrane binding, similarly as
found earlier for caveolins (119, 189). The S-palmitoylation
also facilitates clustering of IFITM3 in the membranes, which is of
potential significance for its antiviral activity (103). In support of the latter, the antiviral capacity was markedly reduced for non-palmitoylated mutant forms of IFITM3 (103, 119). However, S-palmitoylation
did not affect the endolysosomal localization or stability of IFITM3.
Subsequent studies have revealed that the localization and degradation
of murine IFITM3, both shaping its antiviral capacity, are orchestrated
by numerous posttranslational modifications comprising
polyubiquitination, tyrosine phosphorylation by the Src-family kinase
Fyn, and methylation (189, 190). By contrast, S-palmitoylation
alone of the closely related murine IFITM1 endowed it with an antiviral
activity and enhanced stability by preventing proteasomal degradation (55), which indicates diverse effects of this modification on individual IFITM isoforms.
The presented data are only beginning to fill the gap
which existed in our understanding of the role of protein palmitoylation
in innate immune responses. For a long time, it was lagging behind that
on acquired immune responses, in which a plethora of S-palmitoylated
proteins have long been known to be involved. They include receptors
(CD4 and CD8), tyrosine kinases of the Src family, transmembrane adaptor
proteins (e.g., LAT, NTAL, and PAG/Cbp), and α subunits of
heterotrimeric G proteins. Their S-palmitoylation in most cases
targets them to rafts and is a prerequisite for their involvement in the
signaling pathways triggered by immunoreceptors [TCR, B cell receptor
(BCR), and Fcγ and Fcε receptors] crucial for the acquired immune
responses. An association of some components of these signaling pathways
with tetraspanin-enriched domains has also been considered. These
topics are discussed in several earlier reviews (44, 79, 191, 192). It is worth noting that large-scale proteomic analyses of fatty-acylated proteins of T cells (99, 104, 105, 122) and B cells (121),
identifying numerous new palmitoylated proteins, have been published
recently. Further studies will shed light on the possible engagement of
those proteins in acquired immune responses and/or in the cross talk
between the innate and the acquired immune system, in which phagocytic
cells, such as macrophages and dendritic cells, are essential (193).
Concluding Remarks
Protein S-palmitoylation affects their
localization, trafficking, and stability. It has long been known as an
important factor controlling signal transduction by the BCR and TCR
receptors involved in acquired immune responses. It is now becoming
evident that palmitic acid is also a key lipid affecting the diverse
processes at the host–pathogen encounter. Palmitate is a component of
bacterial LPS and lipoproteins; S-palmitoylation of viral, some
bacterial, and numerous host proteins is recognized as a crucial factor
affecting both the virulence of pathogens and the innate immune
reactions of the host. Our understanding of the latter has benefited
greatly from the development of novel methods of detection of this
protein modification. Their application has led to the identification of
numerous proteins involved in the host–pathogen interaction. The
methods have also allowed high-throughput proteomic analysis of
palmitoylation of proteins in infected cells, showing widespread changes
of the host cell palmitoylome. Future studies will tell whether complex
feedback loops comprising palmitoyl acyltransferases and
acylthioesterases, similar to those of kinases and phosphatases carrying
out protein phosphorylation/dephosphorylation, are involved in
controlling protein S-palmitoylation in infected cells. Revealing how the S-palmitoylation
of particular proteins is regulated during the host–pathogen
interactions should allow its modulation to favor the host defense.
Author Contributions
All authors contributed to writing and critically revised the paper.
Abbreviations
17ODYA, 17-octadecynoic acid; ABE, acyl-biotin exchange;
APT, acyl-protein thioesterase; BCR, B cell receptor; FIV, feline
immunodeficiency virus; GPI, glycosylphosphatidylinositol; HA,
hemagglutinin; HCV, hepatitis C virus; HIV-1, human immunodeficiency
virus-1; HSV, herpes simplex virus; IFITM, interferon-induced
transmembrane protein; IFN, interferon; IFNAR, IFNα/β receptor; IL,
interleukin; LPS, lipopolysaccharide; PPT, palmitoyl protein
thioesterase; SILAC, stable isotope labeling by amino acids in cells;
SILAM, stable isotope labeling of mammals; TCR, T cell receptor; TEM8,
tumor endothelial marker 8; TNF, tumor necrosis factor; TNFR, TNF
receptor; TLR, toll-like receptor; VSV, vesicular stomatitis virus;
zDHHC, zinc finger DHHC domain containing.
Inga kommentarer:
Skicka en kommentar