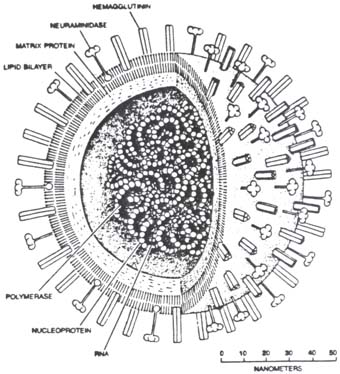
Influenssa A viruksen matrix proteiini M ja M geeni, segmentti 7.
M geeni on bicistroninen. Se tuottaa kahta proteiinia M1 ja M2. M1 on rakenneproteiini ja pääosa viruksen rakenteen suojaproteiineista. M2 on jonikanava .
1. MATRIXPROTEIINI M1 on transkription estäjä.
Matrixproteiini M1 tulee influenssaviruksen geenisegmentistä 7. Uusin tieto mitä siitä näkyy PubMed lähteessä on kaiketi eilispäivältä.
LÄHDE: Huang S, Chen J. Influenza A virus matrix protein 1 interacts with hTFIIIC102-s, a short isoform of the polypeptide 3 subunit of human general transcription factor IIIC. Arch Virol. 2009 Jun 12.
Influenssa A viruksen matrixproteiini 1 (M1) on multifunktionaalinen proteiini, jolla on virionin sisäseinämätehtävän lisäksi myös tärkeää osuutta viruksen replikaation (replication) onnistumisessa, uusien virionien oikeassa kokoontumisessa (assembly) ja vuorautumisessa ja sitten näiden virionien purkaantumisessa itsenäisiksi (budding)
http://www.microbiologybytes.com/virology/3035pics/Orthomyxo3.gif
http://www.schenectadyhistory.org/health/morris/1-100.jpg
Tämä tiedemiesryhmä halusi ottaa selvää, mitkä kaikki isäntäsolun sisällä esiintyvät proteiinit käyvät vuorovaikutukseen tämän virusproteiinin M1 kanssa, kun virus on purkanut sisältönsä isäntäsoluun. He tutkivat yhtätoista(11) itsenäistä proteiinia. Proteiinit olivat ihmissolun cDNA-kirjastosta seulottuja käyttämällä hiivan kahden hybridin tekniikkaa..
Näihin kuuluivat eräs transkriptiotekijä TF ( hTFIIIC102-s protein, encoding a short isoform of the TFIIIC102 subunit of the human TFIIIC transcription factor) Tiedemiehet tutkivat tarkasti mitten viruksen matrixproteiini M1 asettuu interaktioon ihmisen TFIIIC102-s proteiinin kanssa.
Interaktiossa tarvittiin seuraavat kohdat.
M1-proteiinista N-terminaalinen globulaarinen alue( aminohapot 1-164).
http://www2.ujf-grenoble.fr/pharmacie/laboratoires/gdrviro/grouperuigrok/archive/2001/figure2.gif
Ihmisen transkriptiotekijästä hTFIIIC102 viiden( tandem repeat, TR)tetra-tri-kopeptiditoiston (TPR1-5, aminohapot 149-362) alueTämä interaktio vahvistettiin kahdella eri menetelmällä. Lisäksi havaittiin, että HeLa-soluissa samanaikainen ilmenemä hTFIIIC102 transkriptiotekijää ja viruksen esti M1 –matrixproteiinia –translokoitumasta tumaan päin. M1 matrixproteiinia Yhteenvetona tutkijat sanovat, että ihmisen transkriptioproteiini TFIIIC102 voi vaikuttaa influenssaviruksen matrixproteiiniin M1 ja tästä saadaan uusia näkymiä M1-matrisproteiinin rooleihin influenssaviruksen ja isäntäsolun välisessä vuorovaikutuksessa.
2. MATRIX-proteiinista on paljon erikseen myös kirjoitettu, kuten tässä vuonna 2001 (Virus structure group)
http://www2.ujfgrenoble.fr/pharmacie/laboratoires/gdrviro/grouperuigrok/archive/2001/research.htm
LÄHDE: Florence Baudin, Isabelle Petit and rob Ruigrok in collaboration with Winfried Weissenhorn, Steffi Arzt (ESRF, Grenoble), and Wim Burmeister (UJF, Grenoble, visitor Cusack group).
Negatiivisen RNA-juosteen omaavien virusten matrixproteiini M ( tuhkarokon, ebolan, influenssan) omaa aivan välttämätöntä osaa siinä prosessissa, missä uusi virus partikkeli muodostuu infektoituneen solun pinnalla. Yksinkertaistettuna tämän ryhmän viruksilla on kuljetuskomponentit (TRANSPORT componentes) ja replikaatio (REPLICATION) -komponentit. Kuljetuspuolelle kuuluu glykoproteiinitapit joita viruksen lipidipinnalla on kalvorakenteeseen upotettuna( HA, NA) . Nämä proteiinit huolehtivat soluun sisään menosta ( ENTRY) , jotkut virukset myös tuhoavat solureseptoreita siten, että uudet vastamuodostuneet virionit voivat helpommin lähteä ulos solusta. Viruksen kalvo on fysikaalinen este, joka suojaa sisällä olevaa replikaatiokoneistoa ulkoisilta tekijöiltä.
http://s3.images.com/huge.55.278405.JPG
http://edoc.hu-berlin.de/dissertationen/rachakonda-p-sivaramakrishna-2005-02-15/HTML/image002.jpg
REPLIKAATIO-komponentti käsittää viruksen RNA nukleiinihapporakenteen, joka on sitoutuneena stokiometrisella tavalla nukleoproteiiniin NP. Spesifinen viruksen RNA:sta riippuva polymeraasikompleksi on siten sitoutuneena tähän N-RNA kompleksiin. (Sitä sanotaan polymeraasikompleksiksi ja influenssalla siihen kuuluu B2, PB1 ja PA) . Kaiken kaikkiaan totaalinen RNA ja proteiinikomponentit ovat yhteisnimeltään nukleokapsidi NC ja tämä rakenne vastaa viruksen vRNA:n replikaatiosta ja erilaisten mRNA-molekyylien tuotosta heti kun se on päässyt solun sisään ilman että mitään sen enempiä kuljetuskomponentteja ( TRANSPOR components) tarvittaisiin.
VIRUSPARTIKKELIN sisällä matrixproteiinit(M) liimaavat TRANSPORT- ja REPLIKAATIO- komponentteja yhteen . Jotta M pystyisi tekemään sillä tavalla, sen täytyy tehdä kontakti sekä glykoproteiinien sytoplasmisiin päätyihin lipidikalvossa, jolla on spesifinen rakenne (raft) että nukleokapsidin kanssa ja lisäski sisäisesti, sillä se on muodoltaan polymeerinen matrixprotiini, joka vaikuttaa virionin muotoon.
Tämä työryhmä on tutkinut influenssaproteiineja 10 vuotta. ja he ovat myös vertailleet erään toisen työryhmän tietoihin Ebola-viruksen matrixproteiinista (Winfried Weissenhorn)
Influenssaviruksen M-proteiinin spontaaniproteolyysissä havaitaan N-terminaalinen domaani, jossa on aminohappoja 1- 164. Tämä julkaistiin vuonna 2000. Bakteerissa tuotettiin kokomittainen M1 proteiini. Sen N-terminaali domaani ja C-terminaalinen domaani(165-252). Havaittiin, että vain koko pitkä M1 ja sen N-terminaalinen domaani sitoutuivat negatiivisesti varautuneihin liposomeihin ( mikä on yksinkertainen oikean viruksen kalvon vastine kokeissa) ja että vain kokopitkä M1 ja sen C-terminaalinen osa sitoutuivat viraalisen RNP sauvaan.
Kuitenkin vain kokopitkällä M1-molekyylillä oli aktiivisuutta, jolla se esti transkriptiota.
Influenssaviruksen geenistö:
PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS
Toisistaan riippumattomissa kokeissa selviteltiin edelleen asiaa. Koeputkioloissa transkriptio estyi nonspesifisellä M1 proteiinin polymerisaatiolla/aggrekaatiolla RNP-partikkeliin TAI M1 proteiinilla, joka sitoutui toiseen M1-proteiiniin RNP.ssä pikemminkin kuin sitoutumalla NP proteiinin johonkin spesifiseen aktiiviin kohtaan tai polymeraasiin.
Tämä viitaisi siihen, että tämä transkriptiota estävä aktiivisuus, jota kauan jo ollaan kuvattu matrixproteiinin eräänä spesifisena aktiivisuutena, ei näytä olevan spesifinen ollenkaan vaan sivuvaikutus proteiinin luonnollsesta taipumuksesta sitoutua nukleokapsidiin NC ja itse polymerisoitua.
Sen mukaan mitä voimme tästä itse päätellä M-proteiini on aina isoloituna itse viruksesta.
Entä EBOLAN matrixproteiini? VP40, se tuottuu yhtenä domaanina, liukoisena proteiinina, joka destabilisoituessaan tai joutuessaan kontaktiin negatiivissti varautuneiten kalvojen kanssa, avautuu muodostaen kaksi domaania. Sen muutoksen jälkeen VP40 muodostaa oligomeeririnkuloita ja pstyy sitoutumaan kalvoihin. Ei ole vielä testattu pystyykö se myös kiinnittymään nukleokapsideihin. VP40:n kalvoihin sitoutuva aktiivisuus pysyttelee sen C-terminaalisessa domaanissa , kun taas influenssaviruksen matrix-proteiinin M1 vastaava aktiivisuus on N-terminaalisessa domaanissa ja nukleokapsidinkin suhteen käyttäytyy eri tavalla. Vaikuttaa siltä, että näillä kahdella rakenteella on todellakin samoja tehtäviä, mutta ne eivät ole millään muotoa homologisia, mikä näkyy jo niiden atomsesta rakenteesta.
3. VIRUKSEN M-geeni, 8 segmentin geeni tuottaa proteiinit M1 ja M2
M-proteiinien M1 rakenteet ovat virionin päämateriaalia,suojaavaa matrixproteiinia. M2 taas on integralinen membraaniproteiini, jonka tehtvä on tietyssä vaiheessa toimia jonikanavana.
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=249043
Kun on analysoitu M1 ja M2 geenejä ja niiden geenituotteita proteiineja, on havaittu niillä olevan hyvin erilaiset evoluutiotavat. Jos taas verrataan muiden influenssageenien evoluutiotapoihin (esim PB2 ja NP geeneihin), niin M1 ja M2 geenit evoloituvat suhteellisen hitaasti, erityisesti M1 geeni on hidas evoluutiossaan. M1 ja M2 geenituotteet koodautuvat eri lukuraameista (reading frames) mutta toisiaan kattavista (overlapping reading frames) kohdista. Kun näitä tutkittiin, havaittiin, että M1 proteiinit evoloituvat hyvin hitaasti kaikissa linjoissa, kun taas M2 proteiinit osoittavat merkitsevää evoluutiota ihmis-ja sikainfluenssalinjoissa, mutta käytännöllisesti katsoen ei mitään evoluutiota lintuinfluenssalinjoissa.
Evoluutiotahti M1 proteiineissa oli paljon matalampi kuin M2 proteiineissa ja muissa influenssan sisäosan proteiineissa (PB2 ja NP). Samalla M2 proteiineillä taas muihin pintaproteiinehin (esim. H3HA) verrattuna ei ollut niin nopea evoluutio.
Tutkijoiden tulokset osoittavat , että A-influenssaviruksilla bi-cistronisen geenin yhden proteiinin evoluutiolla on merkitystä sen muiden proteiinien evoluutioon
13/06/2009
12:15
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Inga kommentarer:
Skicka en kommentar