EBV viruksen tuma-antigeenin säätelystä
LÄHDE: Berggren Malin (2008, väitöskirja). Regulation and expression of Epstein-Barr virus nuclear antigen 1( EBNA1) in transplant patients and cell culture ISBN 978-91-7433-9
EBV virus kuuluu ihmisen DNA gamma-herpesviruksiin, ja se infektoi yli 90 % ihmiskunnasta. Siis käytännöllisesti katsoen melkein jokaisen- ja on haitaton yleensä. Kun infektio kerran on tullut, se pysyy koko iän siinä isännässä. Infektio voi olla oireeton tai mononukleoosi, joka on itsetään rajoittuva lymfoproliferaatio nuorissa ja aikuisissa. Joskus virus voi aktivoitua uudestaan lyyttisesssä syklissä ja silloin erittyy uutta virusta.
Pysyäkseen sovussa isäntäkehossa virus omaa monimutkaisen sofistisen geenisysteemin ja eri latenssitilat. Sille on eduksi yhteiselo. Siksi useimmilla ei ole mitään oiretta EBV.stä. (Jos tulee lyyttinen sykli, siinä isäntäsolu kuolee, kun virus vapautuu).
Pääasiassa virus on oireeton ja pysyttelee latenssissa tilassaan, jolloin vain muutama sen geeneistä on aktiivina.
(LATENSSIT:
Latenssi III : B-blastisolu. Geeniexpressio EBNA 1-6. LMP1, LMP2.
Latenssi II. Itukeskuksen B-solu EBNA 1, LMP1, LMP2.
Latenssi I. Jakautuva B- muistisolu. EBNA 1.
Latenssi O. Jakaantumaton muistisolu B. Ei geeniexpressiota)
Mutta kun on kyse immuunipuolustukseltaan vaikeammassa tilanteessa olevasta henkilöstä kuten elinsiirtopotilaasta, virus voi joskus aktivoitua uudelleen ja se voi aiheuttaa solun proliferoitumisen, jolloin on riskinä, että kehittyy jokin lymfoproliferatiivinen häiriötila henkilölle, joka on saanut siirrännäisen (PTLD, post transplant lymphoproliferative disorder)
Jakautuvissa soluissa on viruksen replikaatiolle ja erittymiselle tärkeää sen tuma-antigeeni EBNA 1. Se on ainoa virusproteiini, jota esiintyy kaikissa jakautuvissa B-soluissa. Sen takia se on avaintekijä viruksen monitoroinnissa ja mahdollisesti viruksen aktivaatiossa ilmenevien varhaismuutosten havaitsemisessa.
Tässä väitöskirjassa tutkija keskittyi selvittämään EBNA1 esiintymän säätelyä elinsiirtopotilailla.
Keskitytään myös havaittuun IRES kohtaan: EBNA1 geenin translatoitumattomassa 5´alueessa on löydetty IRES, internal ribosome entry site
(IRES on cis- tapaisesti toimiva elementti, joka rekrytoi 40S ribosomaalista kompleksia lähelle AUG- translaation alkavaa kodonia ja täten sallii translaation tapahtuvan CAP- kohdasta riippumatta- eikä täten tarvitse mitään perinteisten aloitustekijöitten kompleksia (Esim: monia eukaryootteja aloitustekijöitä. eIF3, eIF4A, eIF4E, eIF4G, poly A sitova proteiiniPABP yhdistämässä 43S kompleksin, jossa on 40S ribosomi ja mRNA:n CAP- päädyn, jolloin mRNA on suotuisasti translatoitavissa)
Tämä EBNA IRES sallii translaation eteenpäin alavirtaan (downstream), vaikka säännönmukainen CAP-kohdasta riippuvainen translaatio ei toimisi. Tutkijat osoittivat, että EBNA IRES pääaktiviteetti sijaitsi EBV- genomin U-eksonissa. Tämä eksoni IRES- funktioineen näytti olevan hyvin tärkeä EBV-virukselle, koska se oli EBNA 3, 4 ja 6 transkripteissä osana (Nämä ovat EBV latenttiproteiinien joukkoon kuuluvia)
Terveessä väestössä ei löydetä EBNA 1 esiintymää perifeerisessä veressä, mutta sensijaan sitä voi löytää immuunipuolustukseltaan heikommissa. Kun verrattiin elinsiirtopotilaita, joilla oli PTLD oireita, oireettomiin, osoittautui, että PTLD-potilailla oli kolminkertaiset määrät EBNA1- esiintymää veressään. Sen takia EBNA1 esiintymä perifeerisessä veressä voi olla riskin arvioimisessa käypä metodi arvioitaessa elinsiirronjälkeistä vaaraa sairastua lymfoproliferatiiviseen tautiin.
Tutkijat huomasivat myös, että EBNA IRES saattoi joskus deletoitua mRNA:n prosessoiduttua vaihtoehtoisella pilkkoutumisella. Elinsiirtopotilailla esiintyy sekä säännöllisesti että vaihtoehtoisella tavalla pilkkoutunutta EBNA 1 mRNA:ta. Tämä löytö osoittaa epäsuorasti uutta tapaa EBV:n translaation säätymiseen IRES elementin deleetiossa.
LED ja EBV; HUVS ja EBV?
Tutkijat havaitsivat että eräällä keuhkosiirron saaneella potilaalla ei ollut mitään merkkiä PTLD:stä vaikka oli pinttyneesti EBNA1 perifeerisessä veressä. Tällä henkilöllä on HUVS oireyhtymä: Se on harvinainen autoimmuuninen tauti, hypokomplementtinen urtikariatyyppinen vaskuliittioireyhtymä. Tämä HUVS on läheisesti liittynyt LED tautiin, jota on jo aiemmin oletettu liittyväksi EBV-virukseen autoimmuunisten ristiinreagoivien EBNA1 vasta-aineitten takia. Tässä yksittäisessä tapauksessa tutkittiin mahdollisuuksia samantapaiseen assosiatioon HUVS ja EBV viruksen kesken ja tutkijat havaitsivatkin kohonneita vasta-aineita EBNA1 epitoopeille. potilaan seerumissa verrattaessa muihin samanlaisen elinsiirron saaneisiin ja terveisiin verenluovuttajiin. Tämä on ensimmäinen tutkimus EBV- esiintymästä HUVS- taudin yhteydessä ja sen takia lisätutkimukset ovat tarpeen, jotta EBV:n osuus tätä taudissa tulee vahvistetuksi.
Tutkijat katsoivat, mikä on EBNA IRES aktiivisuus lyyttisen induktion aikana. Kaksi reportterivektoria hahmoteltiin- toinen EBNA IRES:llä ja toinen ilman sitä- ja transfektoitiin EBV positiivisiin B-solulinjoihin. jotka olivat kolmessa eri tyypin latenssissa.( Kts. latenssit: )
Transfektoiduissa soluissa indusoitiin lyyttinen vaihe ja EBNA IRES vaikutuksia reportterigeenin ilmenemään tutkittiin proteiini ja RNA- tasoissa.
Tulokset osoittivat että EBNA IRES sisältävissä transfektoiduissa soluissa ilmeni proteiinia 2-3 kertaiset määrät verrattuna soluihin, joissa transfektio suoritettiin ilman EBNA IRES. Nämä tiedot viittaavat siihen, että lyyttisen EBV infektion aikana EBNA IRES omaa aktiivista potentiaalia.
EBV lyyttistä sykliä on vähemmän tutkittu aiemmin, sillä latentit geenit ovat prominentimmat EBV-infektioon liittyvissä maligneissa taudeissa. Mutta kuitenkin on tutkimuksia, jotka ovat osoittaneet lyyttisen EBV-replikaation reaktivaation edeltävän nasofaryngeaalista karsinoomaa, Burkittin lymfoomaa ja Hodginin tautia.
Koska EBV on onkogeeninen ja pystyy välttämään immuunipuolustusta, se on tällä alalla aina mielenkiintoinen virus, jota on tutkittu jo 60 luvulta alkaen.
www.sciencedaily.com
Lähde kertoi 2006-04-04 08:21
EBV tutkimuksista, että viruksen latenttivaihe tarvitsee vain 10 geeniä, kun taas kaikkia muita tarvitaan tuottamaan uutta virusta ja infektoimaan uusia isäntäsoluja soluja. Tämä funktio joka suorittaa vaihteen on ZEEBRA funktio nimeltään ja silloin solu voi tuottaa tuhansia kopioita viruksesta.
Muistiin EBV:stä 08-05-12 03:30 L Bright
torsdag 25 juni 2009
EBV- latenssit ( C Borenström , Theses)
Otan taas muistiinpanoista esiin EBV-viruksen, koska se voi aktivoitua influensaviruksesta, mahdollisesti nimittäin.
EBV viruksesta ( Herpesvirus 4)
LÄHDE. Boreström Cecilia. Regulation of Epstein-Barr virus C promoter by the OriP-EBNA1 complex.
ISBN 978-91-628-7557-2
Asian avainsanoja ovat: Epstein-Barr virus, EBV
EBNA1 (EBV nuclear antigens) Viruksen transkriptiotekijä
NF-Y, Nuclear transcription factor Y
Sp1, Specificity protein 1
Oct-2 ( ihmisen transcriptiotekijä, octamer binding sites)
Geenit jotka kontrolloivat immunoglobuliinien geenien uudelleen järjestäytymistä kautta laajan igh locuksen ja ovat tärkeitä n-lymfosyyttien kehityksen säätelylle.
OCT1 ja 2 transkriptiotekijät ovat tässä tärkeitä eri vaiheissa. ( Asiaa tutkii Stephanie Pritchard-Peter Brodeur 2008)
www.tufts.edu/sackler/mstp/research%20project...
Bright= B-Cell regulator of IgH transcriptionfactor
E2F1 transcriptional regulation
Cp, eräs EBV viruksen vaihteista C promoter.
EBV omaa kyvyn käyttää kolmea eri vaihdetta kolmeen eri latenssiin I, II, III.
http://www.brown.edu/Courses/Bio_160/Projects2000/Herpes/EBV/EBVLC.jpg
Kun EBV ei ole latenteissa vaiheissan, se on lyyttisessä vaiheessa.
Sen kolme promoottoria ovat Qp, Cp ja Wp.
http://www.nature.com/nm/journal/v8/n10/images/nm1002-1086-F1.gif
Ensimmäinen promoottori Qp on käytössä terveillä EBV-viruksen kantajilla, kun virus on kaikkein parhaiten restriktiossaan latenssissaan lepäävissä B-muistisoluissa.
Cp ja Wp promoottorien käyttöä havaitaan, kun alkaa jatkuva proliferaatio (immortalisaatio) ja lymboblastoidien muodostus.
Wp promoottorin käyttö aiheuttaa yhdeksän latentin geenin transkription alkamisen. Nämä ovat EBNA 1-6, joita tuottuu suoraan Wp vaikutuksesta ja EDNA 2, jonka vaikutuksesta alkaa tuottua LMP1, LMP2A ja LMP2B.
oriPI-EDNA1 kompleksi vaikuttaa Wp-Cp vaihteen.
TAUSTAA EBV-viruksesta
EBV virus on yksinomaan ihmisvirus ja se on lymfotrooppinen herpesviruksiin kuuluva ja on infektoinut suurimman osan ihmiskuntaa 90 prosenttisesti kautta maailman.
Primäärinen infektio tapahtuu tavallisesti varhaisina elinvuosina eikä johda mihinkään tunnistettavaan tautiin.
Mutta EBV on myös infektoivan mononukleoosin aiheuttava agenssi.
http://www.terveyskirjasto.fi/terveyskirjasto/tk.koti?p_artikkeli=pot00107
EBV-virukseen liittyy myös pahanlaatuisuuksia
kuten Burkittin lymfooma (BL) http://en.wikipedia.org/wiki/Burkitt%27s_lymphom
Hodgkinin tauti (HD),
http://www.solunetti.fi/se/patologia/hodgkinin_tauti_1/
nenänielun syöpä (nsofaryngeaalinen karsinoma NPC)
ja immunoblastinen lymfoma immuniteetiltään heikentyneissä yksilöissä.
Mutta useimmissa immuniteetiltaan terveissä henkilöissä virus pysyttelee koko elämän latentisti infektoituneissa lepäävissä B-imusoluissa, jotka ovat muistisoluja, aiheuttamatta mitään oireita.
Koeputkessa EBV-pystyy muuttamaan lepääviä B-soluja aktiiveiksi lymfoblasteiksi. Tällaiset aina jakaantuvat solut ilmentävät virusantigeenikirjoa (EBNA-6 ja LMP-1, Latent Membrane proteins ), josta kaikki ovat suoraan osallistumassa immortalisaatioprosessiin. Aivan heti infektion jälkeen Wp ilmentää ( latentteja geenejä) EBNA-2 ja EBNA-5 . Vaihde Cp promoottorille tapahtuu 36 tunnin päästä. Transkriptio tästä ylävirtaan sijaitsevasta C promoottorikohdasta käsin johtaa kaikkien polycistronisesta yksiköstä peräisin olevien EBNA-muotojen transkriptioon siten pilkkoutuen, että geeniluennasta koodautuu erilaisia EBNA proteiineja. EBNA-1 muodostaa multippeleja homodimeerejä, jotka sitoutuvat replikaation latenttisuuden (piilevyyden) alkuperäkohtaan (oriPI), mikä toimii EBNA-1:stä riippuvana C-promoottorikohdan kiihdyttäjänä. Ei ole täysin ymmärretty aiemmin tämän interaktion molekyylitason mekanismia tässä oriPI –EBNA-1 kompleksin ja C-promoottorin välillä.
EBNA-1 ei osoita mitään ilmiselviä entsymaattisia aktiivisuuksia ja arvellaan, että se täyttää tehtävänsä välittämällä interaktioita joillekin spesifisille solussa oleville isäntäproteiineille, joista vain harvoja on luonnehdittu.
Käsillä olevan vaitöskirjan tarkoituksena on ollut identifioida ja luonnehtia tämän makromolekulaarisen kompleksin interaktiopartnereita. Transkriptiotekijän NF-Y ja Sp1 interaktiot ( vuorovaikutukset) C-promoottorin proksimaaliseen alueeseen oli jo aiemmin määritelty tässä laboratoriossa.
(I) Työn ensimmäisessä osassa tutkittiin näitä interaktioita edelleen käyttäen tilapäisiä transfektioita ja varmistettiin, että NF-Y ja Sp1 proteiinit stimuloivat yhdessä C-promoottoria (Cp). Varmistettiin myös, että oriPI-EBNA1 kompleksin aiheuttamassa Cp transaktivaatiossa täytyy välttämättä myös ilmetä samanaikaisesti nämä molemmat proteiinit.NF-Y ja Sp1
(II) Edelleen käytettäessä lymfoblastoidista solulinjaa ( EREB2-5), jossa EBNA-2 funktio on estrogeenisäätöinen, voitiin osoittaa EBNA-2-inaktivaation johtavan NF-Y esiintymään vähenemiseen ja C promoottorin vaimennussäätöön.
Lisäksi kun tiedetään, että lepäävissä soluissa NF-Y ei esiinny ja ottaen huomioon että se vaaditaan kuitenkin C-promoottorin aktivaatioon, tutkijat olivat sitä mieltä, että sen postinfektionelli lisääntymään säätyminen voi osaltaan vaikuttaa Wp- Cp vaihteen primäärissä EBV infektiossa. OriPI sisältää 20 toistodomeenia, jotka sitovat EBNA-1:tä.
Functional Interaction of Nuclear Factor Y and Sp1 is Required for Activation of the Epstein-Barr virus C promoter. Boreström, C., Zetterberg, H., Liff, K., and Rymo, L. Journal of Virology (2003) 77(2): p. 821-9
(II) Työn toisessa osassa tutkijat käyttivät sarjoja oriPI deleetioita ( oriPI-CpCAT reportteriplasmideissa) ja tilapäisiä transfektioita. Sitten he määrittivät EBNA-1 sitovien toistokohtien sen lukumäärän, mikä tarvitaan, jotta Cp välitteinen transaktivaatio olisi tehokas. He pystyivät osoittamaan, että välttämätöntä on 8 tai useamman toistokohdan olemassaolo tälle vaikutukselle. mikä korostaa transaktivaatioprosessin kompleksisuutta
Multiple EBNA1-binding sites within oriPI are required for EBNA1- dependent transactivation of the Epstein-Barr virus C promoter. Zetterberg, H., Boreström, C., Nilsson, T., and Rymo, L. International Journal of Oncology (2004) 25 (3): p 693-6
(III) ja (IV) Näissä työosuuksissa oli tarkoituksena identifioida uusia vaikuttavia partnereita oriPI ja 170 Cp alueissa. Voitiin identifioida kolme uutta proteiini-interaktiota oriPI ja C promoottorin kesken. Nämä kolme proteiinia (Bright (B-cell regulator of Ig H, transkriptiofactor), E2F1 ja Oct-2) näyttivät sitovan molempia sekvenssejä koeputkessa ja in vivo ja tästä aukeni mahdollisuus yhteyden välitykseen oriPI:n ja C promoottorin kesken.
Näiden kolmen proteiinin sitoutumiskohdat kartoitettiin C promoottorissa erääseen lyhyeeseen segmenttiin hyvin lähelle toisiaan. Tämän alueen oli jo aiemmin osoitettu olevan essentielli sekä oriPI:stä riippuville että siitä riippumattomille transkriptionaalisille aktiivisuuksille, mikä viittaa siihen, että interaktio on tärkeä C promoottorin aktiivisuudelle.
Hetkellisin transfektioin osoitettiin , että exogeeninen Oct-2 ja Bright ilmentymä sääti ylös oriPI riippuvaisen C promoottorin aktivaation, kun EBNA-1 ei ollut läsnä.
Lopuksi osoitettiin että endogeeninen Bright ilmenemä korreloi latenssiasteen III malliin, mutta ei latenssiasteitten I ja II ilmentymismalleihin EBV positiivisssa solulinjoissa. Tämä viittaisi edelleen siihen, että Bright ilmenemä ( B-solujen säätelemä IgH transkriptio) olisi tärkeä tekijä C promoottorin transkriptioaktiivisuudessa kehossa.
Functional Interaction of Oct transcription factors with the Family of Repeats in Epstein-Barr virus oriP. Almqvist, J., Zou, J., Linderson, Y., Boreström, C., Altiok, E., Zetterberg, H., Rymo, L., Pettersson, S., and Ernberg, I. Journal of General Virology (2005) 86(5): p. 1261-7
Bright, E2F1 and Oct-2 bind the Epstein-Barr virus C promoter and the oriPI, linking the promoter to the enhancer Boreström, C., Rüetschi, U., and Rymo, L. In manuscript (2008)
Kuvia viruksesta:
www.abc.net.au/.../news/img/health/ebv020306.jpg
www-ermm.cbcu.cam.ac.uk/06010568h.htm
EBV viruksesta ( Herpesvirus 4)
LÄHDE. Boreström Cecilia. Regulation of Epstein-Barr virus C promoter by the OriP-EBNA1 complex.
ISBN 978-91-628-7557-2
Asian avainsanoja ovat: Epstein-Barr virus, EBV
EBNA1 (EBV nuclear antigens) Viruksen transkriptiotekijä
NF-Y, Nuclear transcription factor Y
Sp1, Specificity protein 1
Oct-2 ( ihmisen transcriptiotekijä, octamer binding sites)
Geenit jotka kontrolloivat immunoglobuliinien geenien uudelleen järjestäytymistä kautta laajan igh locuksen ja ovat tärkeitä n-lymfosyyttien kehityksen säätelylle.
OCT1 ja 2 transkriptiotekijät ovat tässä tärkeitä eri vaiheissa. ( Asiaa tutkii Stephanie Pritchard-Peter Brodeur 2008)
www.tufts.edu/sackler/mstp/research%20project...
Bright= B-Cell regulator of IgH transcriptionfactor
E2F1 transcriptional regulation
Cp, eräs EBV viruksen vaihteista C promoter.
EBV omaa kyvyn käyttää kolmea eri vaihdetta kolmeen eri latenssiin I, II, III.
http://www.brown.edu/Courses/Bio_160/Projects2000/Herpes/EBV/EBVLC.jpg
{kind=link}
Kun EBV ei ole latenteissa vaiheissan, se on lyyttisessä vaiheessa.
Sen kolme promoottoria ovat Qp, Cp ja Wp.
http://www.nature.com/nm/journal/v8/n10/images/nm1002-1086-F1.gif
{kind=link}
Ensimmäinen promoottori Qp on käytössä terveillä EBV-viruksen kantajilla, kun virus on kaikkein parhaiten restriktiossaan latenssissaan lepäävissä B-muistisoluissa.
Cp ja Wp promoottorien käyttöä havaitaan, kun alkaa jatkuva proliferaatio (immortalisaatio) ja lymboblastoidien muodostus.
Wp promoottorin käyttö aiheuttaa yhdeksän latentin geenin transkription alkamisen. Nämä ovat EBNA 1-6, joita tuottuu suoraan Wp vaikutuksesta ja EDNA 2, jonka vaikutuksesta alkaa tuottua LMP1, LMP2A ja LMP2B.
oriPI-EDNA1 kompleksi vaikuttaa Wp-Cp vaihteen.
TAUSTAA EBV-viruksesta
EBV virus on yksinomaan ihmisvirus ja se on lymfotrooppinen herpesviruksiin kuuluva ja on infektoinut suurimman osan ihmiskuntaa 90 prosenttisesti kautta maailman.
Primäärinen infektio tapahtuu tavallisesti varhaisina elinvuosina eikä johda mihinkään tunnistettavaan tautiin.
Mutta EBV on myös infektoivan mononukleoosin aiheuttava agenssi.
http://www.terveyskirjasto.fi/terveyskirjasto/tk.koti?p_artikkeli=pot00107
EBV-virukseen liittyy myös pahanlaatuisuuksia
kuten Burkittin lymfooma (BL) http://en.wikipedia.org/wiki/Burkitt%27s_lymphom
Hodgkinin tauti (HD),
http://www.solunetti.fi/se/patologia/hodgkinin_tauti_1/
nenänielun syöpä (nsofaryngeaalinen karsinoma NPC)
ja immunoblastinen lymfoma immuniteetiltään heikentyneissä yksilöissä.
Mutta useimmissa immuniteetiltaan terveissä henkilöissä virus pysyttelee koko elämän latentisti infektoituneissa lepäävissä B-imusoluissa, jotka ovat muistisoluja, aiheuttamatta mitään oireita.
Koeputkessa EBV-pystyy muuttamaan lepääviä B-soluja aktiiveiksi lymfoblasteiksi. Tällaiset aina jakaantuvat solut ilmentävät virusantigeenikirjoa (EBNA-6 ja LMP-1, Latent Membrane proteins ), josta kaikki ovat suoraan osallistumassa immortalisaatioprosessiin. Aivan heti infektion jälkeen Wp ilmentää ( latentteja geenejä) EBNA-2 ja EBNA-5 . Vaihde Cp promoottorille tapahtuu 36 tunnin päästä. Transkriptio tästä ylävirtaan sijaitsevasta C promoottorikohdasta käsin johtaa kaikkien polycistronisesta yksiköstä peräisin olevien EBNA-muotojen transkriptioon siten pilkkoutuen, että geeniluennasta koodautuu erilaisia EBNA proteiineja. EBNA-1 muodostaa multippeleja homodimeerejä, jotka sitoutuvat replikaation latenttisuuden (piilevyyden) alkuperäkohtaan (oriPI), mikä toimii EBNA-1:stä riippuvana C-promoottorikohdan kiihdyttäjänä. Ei ole täysin ymmärretty aiemmin tämän interaktion molekyylitason mekanismia tässä oriPI –EBNA-1 kompleksin ja C-promoottorin välillä.
EBNA-1 ei osoita mitään ilmiselviä entsymaattisia aktiivisuuksia ja arvellaan, että se täyttää tehtävänsä välittämällä interaktioita joillekin spesifisille solussa oleville isäntäproteiineille, joista vain harvoja on luonnehdittu.
Käsillä olevan vaitöskirjan tarkoituksena on ollut identifioida ja luonnehtia tämän makromolekulaarisen kompleksin interaktiopartnereita. Transkriptiotekijän NF-Y ja Sp1 interaktiot ( vuorovaikutukset) C-promoottorin proksimaaliseen alueeseen oli jo aiemmin määritelty tässä laboratoriossa.
(I) Työn ensimmäisessä osassa tutkittiin näitä interaktioita edelleen käyttäen tilapäisiä transfektioita ja varmistettiin, että NF-Y ja Sp1 proteiinit stimuloivat yhdessä C-promoottoria (Cp). Varmistettiin myös, että oriPI-EBNA1 kompleksin aiheuttamassa Cp transaktivaatiossa täytyy välttämättä myös ilmetä samanaikaisesti nämä molemmat proteiinit.NF-Y ja Sp1
(II) Edelleen käytettäessä lymfoblastoidista solulinjaa ( EREB2-5), jossa EBNA-2 funktio on estrogeenisäätöinen, voitiin osoittaa EBNA-2-inaktivaation johtavan NF-Y esiintymään vähenemiseen ja C promoottorin vaimennussäätöön.
Lisäksi kun tiedetään, että lepäävissä soluissa NF-Y ei esiinny ja ottaen huomioon että se vaaditaan kuitenkin C-promoottorin aktivaatioon, tutkijat olivat sitä mieltä, että sen postinfektionelli lisääntymään säätyminen voi osaltaan vaikuttaa Wp- Cp vaihteen primäärissä EBV infektiossa. OriPI sisältää 20 toistodomeenia, jotka sitovat EBNA-1:tä.
Functional Interaction of Nuclear Factor Y and Sp1 is Required for Activation of the Epstein-Barr virus C promoter. Boreström, C., Zetterberg, H., Liff, K., and Rymo, L. Journal of Virology (2003) 77(2): p. 821-9
(II) Työn toisessa osassa tutkijat käyttivät sarjoja oriPI deleetioita ( oriPI-CpCAT reportteriplasmideissa) ja tilapäisiä transfektioita. Sitten he määrittivät EBNA-1 sitovien toistokohtien sen lukumäärän, mikä tarvitaan, jotta Cp välitteinen transaktivaatio olisi tehokas. He pystyivät osoittamaan, että välttämätöntä on 8 tai useamman toistokohdan olemassaolo tälle vaikutukselle. mikä korostaa transaktivaatioprosessin kompleksisuutta
Multiple EBNA1-binding sites within oriPI are required for EBNA1- dependent transactivation of the Epstein-Barr virus C promoter. Zetterberg, H., Boreström, C., Nilsson, T., and Rymo, L. International Journal of Oncology (2004) 25 (3): p 693-6
(III) ja (IV) Näissä työosuuksissa oli tarkoituksena identifioida uusia vaikuttavia partnereita oriPI ja 170 Cp alueissa. Voitiin identifioida kolme uutta proteiini-interaktiota oriPI ja C promoottorin kesken. Nämä kolme proteiinia (Bright (B-cell regulator of Ig H, transkriptiofactor), E2F1 ja Oct-2) näyttivät sitovan molempia sekvenssejä koeputkessa ja in vivo ja tästä aukeni mahdollisuus yhteyden välitykseen oriPI:n ja C promoottorin kesken.
Näiden kolmen proteiinin sitoutumiskohdat kartoitettiin C promoottorissa erääseen lyhyeeseen segmenttiin hyvin lähelle toisiaan. Tämän alueen oli jo aiemmin osoitettu olevan essentielli sekä oriPI:stä riippuville että siitä riippumattomille transkriptionaalisille aktiivisuuksille, mikä viittaa siihen, että interaktio on tärkeä C promoottorin aktiivisuudelle.
Hetkellisin transfektioin osoitettiin , että exogeeninen Oct-2 ja Bright ilmentymä sääti ylös oriPI riippuvaisen C promoottorin aktivaation, kun EBNA-1 ei ollut läsnä.
Lopuksi osoitettiin että endogeeninen Bright ilmenemä korreloi latenssiasteen III malliin, mutta ei latenssiasteitten I ja II ilmentymismalleihin EBV positiivisssa solulinjoissa. Tämä viittaisi edelleen siihen, että Bright ilmenemä ( B-solujen säätelemä IgH transkriptio) olisi tärkeä tekijä C promoottorin transkriptioaktiivisuudessa kehossa.
Functional Interaction of Oct transcription factors with the Family of Repeats in Epstein-Barr virus oriP. Almqvist, J., Zou, J., Linderson, Y., Boreström, C., Altiok, E., Zetterberg, H., Rymo, L., Pettersson, S., and Ernberg, I. Journal of General Virology (2005) 86(5): p. 1261-7
Bright, E2F1 and Oct-2 bind the Epstein-Barr virus C promoter and the oriPI, linking the promoter to the enhancer Boreström, C., Rüetschi, U., and Rymo, L. In manuscript (2008)
Kuvia viruksesta:
www.abc.net.au/.../news/img/health/ebv020306.jpg
www-ermm.cbcu.cam.ac.uk/06010568h.htm
onsdag 17 juni 2009
Pandeminen A/H1N1 , lääkkeet ja rokotekehittely
http://leabright.wordpress.com/2009/06/16/1-swedish-medical-journal-about-pandemic-flu/
http://leabright.wordpress.com/2009/06/16/2-swedish-medical-journal-about-pandemic-flu-and-vaccin/
http://leabright.wordpress.com/2009/06/16/3-swedish-medical-journal-about-pandemic-vaccin/
http://leabright.wordpress.com/2009/06/17/5-swedish-medical-journal-about-antivirasl-in-pandemic-ah1n1/
http://leabright.wordpress.com/2009/06/17/6swedish-medical-journal-about-resistance-against-antivirals/
http://leabright.wordpress.com/2009/06/16/2-swedish-medical-journal-about-pandemic-flu-and-vaccin/
http://leabright.wordpress.com/2009/06/16/3-swedish-medical-journal-about-pandemic-vaccin/
http://leabright.wordpress.com/2009/06/17/5-swedish-medical-journal-about-antivirasl-in-pandemic-ah1n1/
http://leabright.wordpress.com/2009/06/17/6swedish-medical-journal-about-resistance-against-antivirals/
söndag 14 juni 2009
Influenssaviruksen glykosylaatioista (HA)
Influenssavirus glykosyloi omat pintaproteiininsa ihmisen glykosylaaseilla solun sisällä ollen.
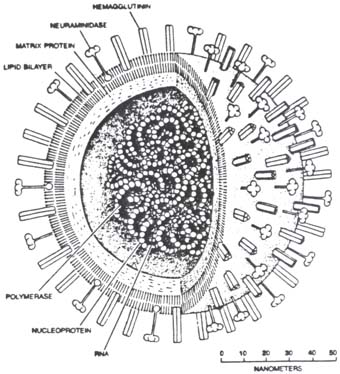
(Influenssaviruksen geenit PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS)
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1161681
Pohdittaa tuo virusten proteiinien tapa hankkia glykosylaationsa. Olisi luultavaa että seriinin (S) mutaatio asparagiiniksi (N) tekisi viruksen glykosylaatiomahdollisuuksista ehkä pahemman laatuisia ja samalla glykosylaatiotyyppi olennaisesti muuttuu.Se vaikuttaa invasiivisuuteen ja virulenssiin, aivan kuten onkologian alalla. Mitähän ihmisentsyymiä influenssavirus käyttää proteiiniensa glykosyloimiseen?
GLYKOFOORUMIN uutisia pitää katsoa jos tekee muistiinpanot HA ja NA virusproteiineista, jotka tarvitsevat glykosylaatiota.
http://www.glycoforum.gr.jp/science/glycomicrobiology/GM04/GM04E.html
(Influenssaviruksen geenit PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS)
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1161681
Pohdittaa tuo virusten proteiinien tapa hankkia glykosylaationsa. Olisi luultavaa että seriinin (S) mutaatio asparagiiniksi (N) tekisi viruksen glykosylaatiomahdollisuuksista ehkä pahemman laatuisia ja samalla glykosylaatiotyyppi olennaisesti muuttuu.Se vaikuttaa invasiivisuuteen ja virulenssiin, aivan kuten onkologian alalla. Mitähän ihmisentsyymiä influenssavirus käyttää proteiiniensa glykosyloimiseen?
GLYKOFOORUMIN uutisia pitää katsoa jos tekee muistiinpanot HA ja NA virusproteiineista, jotka tarvitsevat glykosylaatiota.
http://www.glycoforum.gr.jp/science/glycomicrobiology/GM04/GM04E.html
Influenssavirus: N40, PB1, PB1-F2
Aiemmin on arveltu, että kahdeksan influensaviruksen genomisegmenttiä tuottaa yksitoista proteiinituotetta, mutta nyt siihen on lisätty kahdestoista joukkoon N40 nimeltään. Mistä se muodostuu?
Tämä onkin monimutkainen seikka, sillä segmentti 2 voi tuottaa joko PB1 proteiinin tai toisella luennalla PB1-F2 proteiinin ja tähän on nyt lisätty kolmas tuote N40 seuraavasti.
LÄHDE: Wise HM, Foeglein A et al. A complicated message: identification of a novel PB1-related protein translated from influenza A segment 2 mRNA. J Virol. 2009 Jun 3. Division of Virology, Department of Pathology, University of Cambridge, UK.
Suomennosta:
Influenssa A viruksen segmentin( geenin) 2 tiedetään koodaavan kahta polypeptidiä joilla on toisiaan kattava ORF (Open Reading Frames).
PB1 on polymeraasi nimeltään.
PB1-F2 on proapotoottinen virulenssitekijä.
Nyt tutkijat osoittivat kolmannen polypeptidin syntetisoituvatn PB1-mRNA:sta eri AUG starttikodonia käyttämällä. PB1 kodoni 40 johtaa translaatiota, josta tuottuu N-terminaalisesti lyhentynyt versio polypeptidiä ja sitä sanotaan N40:ksi. Siltä puuttuu trankrisptaasifunktio, mutta siitä huolimatta se käy interaktioon PB2-proteiinin ja polymeraasikompleksin kanssa solumiljöössä.
On tärkeä huomata että N40, PB1-F2 ja PB1 polypeptidien expressio on keskinäisesti toisistaan riippuvia. Tietyt mutaatiot, joita on käytetty poistamaan PB1-F2 tuotanto , vaikuttavat N40 peptidin kertymisiin.
Jos PB1-F2:n AUG (starttikohta) poistetaan, säätyy N40 peptidin synteesi ylös.
Jos taas PB1-F2 lyhennetään kodonin 8 jälkeen ( sitä seuraavalla M401 muutoksella PB1 rakenteeseen) , pyyhkiytyy N40 expressio pois.
Sellainen virus jolta puuttuu polypeptidit N40 ja PB1-F2, replikoituu normaalisti.
Kuitenkin ne virukset, jotka eivät ilmennä N40 polypeptidiä, mutta säilyttävät intaktin PB1-F2 geeninsä, ilmentävät ylimäärin PB1 proteiinia infektion varhaisvaiheessa ja replikoituvat hitaasti kudoksessa.
Täten influenssa A viruksen proteomissa on kahdestoistakin translaatiotuote, joka (samoin kuin PB1-F2) ei ole välttämätön viruksen elinkyvylle, mutta jonka menettäminen varsinkin sen geenitaustassa, on haitallinen viruksen replikaatiolle
Tämä onkin monimutkainen seikka, sillä segmentti 2 voi tuottaa joko PB1 proteiinin tai toisella luennalla PB1-F2 proteiinin ja tähän on nyt lisätty kolmas tuote N40 seuraavasti.
LÄHDE: Wise HM, Foeglein A et al. A complicated message: identification of a novel PB1-related protein translated from influenza A segment 2 mRNA. J Virol. 2009 Jun 3. Division of Virology, Department of Pathology, University of Cambridge, UK.
Suomennosta:
Influenssa A viruksen segmentin( geenin) 2 tiedetään koodaavan kahta polypeptidiä joilla on toisiaan kattava ORF (Open Reading Frames).
PB1 on polymeraasi nimeltään.
PB1-F2 on proapotoottinen virulenssitekijä.
Nyt tutkijat osoittivat kolmannen polypeptidin syntetisoituvatn PB1-mRNA:sta eri AUG starttikodonia käyttämällä. PB1 kodoni 40 johtaa translaatiota, josta tuottuu N-terminaalisesti lyhentynyt versio polypeptidiä ja sitä sanotaan N40:ksi. Siltä puuttuu trankrisptaasifunktio, mutta siitä huolimatta se käy interaktioon PB2-proteiinin ja polymeraasikompleksin kanssa solumiljöössä.
On tärkeä huomata että N40, PB1-F2 ja PB1 polypeptidien expressio on keskinäisesti toisistaan riippuvia. Tietyt mutaatiot, joita on käytetty poistamaan PB1-F2 tuotanto , vaikuttavat N40 peptidin kertymisiin.
Jos PB1-F2:n AUG (starttikohta) poistetaan, säätyy N40 peptidin synteesi ylös.
Jos taas PB1-F2 lyhennetään kodonin 8 jälkeen ( sitä seuraavalla M401 muutoksella PB1 rakenteeseen) , pyyhkiytyy N40 expressio pois.
Sellainen virus jolta puuttuu polypeptidit N40 ja PB1-F2, replikoituu normaalisti.
Kuitenkin ne virukset, jotka eivät ilmennä N40 polypeptidiä, mutta säilyttävät intaktin PB1-F2 geeninsä, ilmentävät ylimäärin PB1 proteiinia infektion varhaisvaiheessa ja replikoituvat hitaasti kudoksessa.
Täten influenssa A viruksen proteomissa on kahdestoistakin translaatiotuote, joka (samoin kuin PB1-F2) ei ole välttämätön viruksen elinkyvylle, mutta jonka menettäminen varsinkin sen geenitaustassa, on haitallinen viruksen replikaatiolle
NP, influenssaviruksen nukleoproteiini ja NLS (Segm.5)
5.geenisegmentti ja sen koodaama proteiini NP. Nukleoproteiini.
Influenssaviruksen nukleoproteiini NP on kuin viruksen vene.
Nukleoproteiinin varassa viruksen replikatiivinen koneisto pääsee lipumaan tumaan sen jälkeen, kun viruskapseli on tyhjentänyt nämä sisärakenteensa työkalut solulimaan.
Nukleoproteiiniveneen varassa sitten työkalulaitteisto menee tumaan operoimaan ja käyttää myös uusista nukleoproteiineista tehtyä nukleoproteiinialustaa kuin lauttana, johon uudet virukset sitten rakennetaan ja palautetaan sytoplasman nesteen läpi takaisin uusiin virionpartikkeleisiin.
Kuten yleensäkin veneessä tiedetään liikkuminen on polarisoitua, kokka edellä. Samalla tavalla nukleokapsidin vRNP partikkelit ovat tilanteeseen polarisoituja kulkiessaan eri suuntiin, ensin kohti tumaa ja lopuksi uutena partikkelisukupolvena pois tumasta kohti solun periferiaa valmiina odottaviin virionikuoriin, jotka jostain syystä ovat kypsiä sulkeutumaan ja lähtemään irti solun hukkuvasta laivasta, kun kaikkia 8 segmenttiä, viruksen tiimin jäsentä on saapunut paikalle.
Virion taas toimii kuin sukelluskello näille pikkupartikkeleille, viruksen genomiselle tiimille . Ne eivät tuota replikoitumista virionin sisällä, vaan odottavat mahdollisuutta päästä taas uuteen soluun ja seilaamaan sytoplasman läpi isäntätumaan. Segmentit ovat kuin ”veneet vedettynä kuiville” ja kumolleen talveksi, kun ne ovat virionin sisällä inaktiivissa tilassa ahtaasti nukleokapsidirakenteessaan.
http://edoc.hu-berlin.de/dissertationen/rachakonda-p-sivaramakrishna-2005-02-15/HTML/image002.jpg
Kuva osoittaa, miten erittäin hyvin organisoitua ja ohjelmoituja ovat influenssaviruksen toiminnot.
Tästä käsittää, että nukleoproteiinilla on suuri merkitys tapahtumien alustana, telakkana, minkä varassa koko viruksen siirtyileminen, lisääntyminen solussa ja säilyminen virioneissa tapahtuu. Tuollainen negatiivinen RNA säie ei ole millään muotoa elinkykyinen ilman suurta suojaavaa systeemiä.
Miten tiede asian sanoo?
LÄHDE: Portela A, Digard P.
The influenza virus nucleoprotein: a multifunctional RNA-binding protein pivotal to virus replication. J Gen Virol. 2002 Apr;83(Pt 4):723-34. División de Productos Biológicos y Biotecnología, Agencia Española del Medicamento, Majadahonda 28220, Madrid, Spain.
(Suomennosta)
KAIKKI negatiivisen (-) RNA-genomin omaavat virukset koodaavat yksinkertaista RNA-säiettä sitovaa nukleoproteiinia (NP)
NUKLEOPROTEIININ PRIMÄÄRINÄ FUNKTIONA on tehdä viruksen genomille nukleokapsidi (NC), joka sitten pystyy RNA:n transkriptioon, replikaatioon ja pakkaustoimiin.
Tutkijat halusivat tässä katsauksessaan antaa kuvaa influenssavirukseta sen NP-proteiinin avulla, miten tämä molekyyli on paljon enemmän kuin vain rakenteellinen RNA-materiaalia sitova proteiini. Se on lisäksi avainasemassa oleva adaptorimolekyyli, joka sopeuttaa viruksen ja isäntäsolun väliset prosessit toisiinsa.
Se voi suorittaa näitä tehtäviänsä, koska sillä on kykyä asettua interaktioon useitten virusperäisten ja isäntäsoluperäisten makromolekyylien kanssa, joihin kuuluu
Tutkijat tarkastelivat työssään näitten mainittujen aktiviteettien olemassaoloa ja niitten mahdollista osuutta virusgenomin transkriptiossa, replikaatiossa ja solun sisäisessä kuljetuksessa.
TOISESSA LÄHTEESSÄ kirjoitetaan minkälaisella “kompassilla” nukleoproteiini voi ohjata täsmällisesti virusainesta oikeisiin suuntiin.
LÄHDE 2. Wu WW, Sun YH, Panté N. Nuclear import of influenza A viral ribonucleoprotein complexes is mediated by two nuclear localization sequences (NLS) on viral nucleoprotein. Virol J. 2007 Jun 4;4:49 Department of Zoology, University of British Columbia, Vancouver, BC, Canada.
(Suomennosta)
Influenssavirus replikoituu ( tekee lisää genomista materiaaliaan) isäntäsolun tumassa.
Jotta infektoitumista ylipäätänä tapahtuisi, viruksen on mentävä solun TUMAAN asti.
Viruksen nukleoproteiinin(NP) useat kopiot (NP monomeerit) auttavat pakkaamaan viruksen kahdeksan eri vRNA säiettä yksittäin ribonukleoproteiini-partikkeleiksi (vRNP). Kun jokainen pakkaus on valmis, ne ovat isoja sauvamaisia komplekseja, joiden ytimen muodostaa tämä NP, nukleoproteiini ja viruksen RNA on kietoutunut helixmäisesti sen ympäri.
Nämä vRNP-sauvat ovat se osa koko virusta, jonka on päästävä, uiskenneltava, tuman sisään. Sitä tarkoitusta varten niillä täytyy olla jokin kompassin tapainen orientoituminen. Sitä sanotaan tumaan lokalisoivaksi jaksoksi rakenteessa ( nuclear localization sequences, NLS). Sellaisen jakson täytyy olla jollain tavalla vRNP sauvan pinnalla.
Oletetaan että NP sisältää kaksi NLS-kohtaa, joista toinen on N-terminaalinen (NLS1) ja toinen on keskellä (NLS2) proteiinia .
On havaittu että rekombinanteissa NP-molekyyleissä nämä NP proteiinin NLS-kohdat välittävät sauvan sisään menoa tumaan (import). Tutkijoitten oli vielä selvitettävä mikä NLS influensaviruksen vRNP-sauvassa määräsi tumaan sisäänmenoa. Tästä he tekivät ylläolevan työnsä.
Havaittiin, että molemmat NLS-kohdat toimivat itsenäisesti. Jos kohdat blokeerattiin, vRNP tuskin ollenkaan pääsi tumaan. Havaittiin että NLS1 oli päätekijä vRNP sauvan tumaan pääsyssä ja
jos kaksi kohtaa oli vaikuttamassa, tumaan pääsy oli parempi.
Nämä NLS 1 ja NLS 2 kohdat (”virusvenettä” orientoivat kohdat) ovat tiedemiesten tiedossa.
Influenssaviruksen nukleoproteiini NP on kuin viruksen vene.
Nukleoproteiinin varassa viruksen replikatiivinen koneisto pääsee lipumaan tumaan sen jälkeen, kun viruskapseli on tyhjentänyt nämä sisärakenteensa työkalut solulimaan.
Nukleoproteiiniveneen varassa sitten työkalulaitteisto menee tumaan operoimaan ja käyttää myös uusista nukleoproteiineista tehtyä nukleoproteiinialustaa kuin lauttana, johon uudet virukset sitten rakennetaan ja palautetaan sytoplasman nesteen läpi takaisin uusiin virionpartikkeleisiin.
Kuten yleensäkin veneessä tiedetään liikkuminen on polarisoitua, kokka edellä. Samalla tavalla nukleokapsidin vRNP partikkelit ovat tilanteeseen polarisoituja kulkiessaan eri suuntiin, ensin kohti tumaa ja lopuksi uutena partikkelisukupolvena pois tumasta kohti solun periferiaa valmiina odottaviin virionikuoriin, jotka jostain syystä ovat kypsiä sulkeutumaan ja lähtemään irti solun hukkuvasta laivasta, kun kaikkia 8 segmenttiä, viruksen tiimin jäsentä on saapunut paikalle.
Virion taas toimii kuin sukelluskello näille pikkupartikkeleille, viruksen genomiselle tiimille . Ne eivät tuota replikoitumista virionin sisällä, vaan odottavat mahdollisuutta päästä taas uuteen soluun ja seilaamaan sytoplasman läpi isäntätumaan. Segmentit ovat kuin ”veneet vedettynä kuiville” ja kumolleen talveksi, kun ne ovat virionin sisällä inaktiivissa tilassa ahtaasti nukleokapsidirakenteessaan.
http://edoc.hu-berlin.de/dissertationen/rachakonda-p-sivaramakrishna-2005-02-15/HTML/image002.jpg
{kind=link}
Kuva osoittaa, miten erittäin hyvin organisoitua ja ohjelmoituja ovat influenssaviruksen toiminnot.
Tästä käsittää, että nukleoproteiinilla on suuri merkitys tapahtumien alustana, telakkana, minkä varassa koko viruksen siirtyileminen, lisääntyminen solussa ja säilyminen virioneissa tapahtuu. Tuollainen negatiivinen RNA säie ei ole millään muotoa elinkykyinen ilman suurta suojaavaa systeemiä.
Miten tiede asian sanoo?
LÄHDE: Portela A, Digard P.
The influenza virus nucleoprotein: a multifunctional RNA-binding protein pivotal to virus replication. J Gen Virol. 2002 Apr;83(Pt 4):723-34. División de Productos Biológicos y Biotecnología, Agencia Española del Medicamento, Majadahonda 28220, Madrid, Spain.
(Suomennosta)
KAIKKI negatiivisen (-) RNA-genomin omaavat virukset koodaavat yksinkertaista RNA-säiettä sitovaa nukleoproteiinia (NP)
NUKLEOPROTEIININ PRIMÄÄRINÄ FUNKTIONA on tehdä viruksen genomille nukleokapsidi (NC), joka sitten pystyy RNA:n transkriptioon, replikaatioon ja pakkaustoimiin.
Tutkijat halusivat tässä katsauksessaan antaa kuvaa influenssavirukseta sen NP-proteiinin avulla, miten tämä molekyyli on paljon enemmän kuin vain rakenteellinen RNA-materiaalia sitova proteiini. Se on lisäksi avainasemassa oleva adaptorimolekyyli, joka sopeuttaa viruksen ja isäntäsolun väliset prosessit toisiinsa.
Se voi suorittaa näitä tehtäviänsä, koska sillä on kykyä asettua interaktioon useitten virusperäisten ja isäntäsoluperäisten makromolekyylien kanssa, joihin kuuluu
- itse RNA,
- kaksi virusperäisestä RNA:sta riippuvaa RNA-polymeraasialayksikköä
- matrixproteiini M.
- Lisäksi NP asettuu interaktioon solun polypeptidien kanssa ja niihin kuuluu
- aktiini
- solutuman import ja export- laitteiston komponentit
- ja tuman RNA-helikaasit.
Tutkijat tarkastelivat työssään näitten mainittujen aktiviteettien olemassaoloa ja niitten mahdollista osuutta virusgenomin transkriptiossa, replikaatiossa ja solun sisäisessä kuljetuksessa.
TOISESSA LÄHTEESSÄ kirjoitetaan minkälaisella “kompassilla” nukleoproteiini voi ohjata täsmällisesti virusainesta oikeisiin suuntiin.
LÄHDE 2. Wu WW, Sun YH, Panté N. Nuclear import of influenza A viral ribonucleoprotein complexes is mediated by two nuclear localization sequences (NLS) on viral nucleoprotein. Virol J. 2007 Jun 4;4:49 Department of Zoology, University of British Columbia, Vancouver, BC, Canada.
(Suomennosta)
Influenssavirus replikoituu ( tekee lisää genomista materiaaliaan) isäntäsolun tumassa.
Jotta infektoitumista ylipäätänä tapahtuisi, viruksen on mentävä solun TUMAAN asti.
Viruksen nukleoproteiinin(NP) useat kopiot (NP monomeerit) auttavat pakkaamaan viruksen kahdeksan eri vRNA säiettä yksittäin ribonukleoproteiini-partikkeleiksi (vRNP). Kun jokainen pakkaus on valmis, ne ovat isoja sauvamaisia komplekseja, joiden ytimen muodostaa tämä NP, nukleoproteiini ja viruksen RNA on kietoutunut helixmäisesti sen ympäri.
Nämä vRNP-sauvat ovat se osa koko virusta, jonka on päästävä, uiskenneltava, tuman sisään. Sitä tarkoitusta varten niillä täytyy olla jokin kompassin tapainen orientoituminen. Sitä sanotaan tumaan lokalisoivaksi jaksoksi rakenteessa ( nuclear localization sequences, NLS). Sellaisen jakson täytyy olla jollain tavalla vRNP sauvan pinnalla.
Oletetaan että NP sisältää kaksi NLS-kohtaa, joista toinen on N-terminaalinen (NLS1) ja toinen on keskellä (NLS2) proteiinia .
On havaittu että rekombinanteissa NP-molekyyleissä nämä NP proteiinin NLS-kohdat välittävät sauvan sisään menoa tumaan (import). Tutkijoitten oli vielä selvitettävä mikä NLS influensaviruksen vRNP-sauvassa määräsi tumaan sisäänmenoa. Tästä he tekivät ylläolevan työnsä.
Havaittiin, että molemmat NLS-kohdat toimivat itsenäisesti. Jos kohdat blokeerattiin, vRNP tuskin ollenkaan pääsi tumaan. Havaittiin että NLS1 oli päätekijä vRNP sauvan tumaan pääsyssä ja
jos kaksi kohtaa oli vaikuttamassa, tumaan pääsy oli parempi.
Nämä NLS 1 ja NLS 2 kohdat (”virusvenettä” orientoivat kohdat) ovat tiedemiesten tiedossa.
NP, influenssaviruksen nukleoproteiini ja ihmisen NF
Miten ihmsen solu voisi puolustautua viruksen nukleoproteiinia NP vastaan?
Yksi seikka ontumatekijä NF90.
Mikä on ihmisen solun tumatekijä NF90?
Tumatekijä NF90 kuuluu tunnettuihin säätelijämolekyyleihin, jotka sitoutuvat antigeenivasteen tunnistuselementtiin ARRE-2 ja stimuloivat transkriptiota ja translaatiota normaalisolussa.
Mutta uutena tietona on, että tumatekijä NF90 voi myös negatiiviseti säädellä influenssaviruksen replikaatiota tekemällä vuorovaikutuksen influenssaviruksen nukleoproteiiniin NP, jos sitä esiintyy tumassa.
TUORE LÄHDE: http://www.citeulike.org/user/friendpine/article/4768934
Wang P, Song W et al. Nuclear factor 90, NF90, negatively regulates influenza virus replication by interacting with viral nucleoprotein. J Virol. 2009 Jun 3. [Epub ahead of print]
ISÄNTÄKEHON TEKIJÖIDEN ja VIRUSREPLIKAATION interaktiot ovat tärkeitä isäntäkehon vasteessa influenssavirukseen ja sen replikaation säätelyssä.
SOLUPROTEIINIA, jonka nimi on tumatekijä 90 (nuclear factor 90, NF90) , on löydetty ihmissoluista, joissa on esiintynyt ohimenevästi viruksen nukleoproteiinia NP ja tämä NF90 on löydetty samassa yhteydessä kun on puhdistettu viruksen H5N1 nukleoproteiinia NP käyttäen menetelmänä Mass Spectrometry (MALDI-TOF MS) analyysiä.
Koeputkessa NF90 ja NP-proteiinit saostuivat samanaikaisesti eräissä viljellyissä soluissa tai niitä esiintyi yksinään bakteerisoluissa tai kudosviljelmäsoluissa vastaavasti. Tämä löytö vahvisti, että NF90 ja NP välillä tapahtuu interaktiota itsenäisesti muista viruspartikkelin RNP proteiineista riippumatta.
Jos viruksen NP proteiinissa oli C-terminaalinen mutaatio (F412A) , interaktiota soluproteiiniin NF90 ei tapahtunut, joten tästä pääteltiin, että viruksen NP vaatii C- terminaalia sitoakseen NF90-proteiinia.
RNaasi V-käsittely ei asettanut esteitä NP ja NF90-proteiinien yhteiselle saostumiselle, mikä demonstroi, että niiden interaktio ei ollut riippuvainen RNA:sta.
Jos siRNA:lla tehdyn poistogeenisen NF90 vajeisen solun interaktioita tutkittiin, havaittiin, että siinä virusten polymeraasikompleksin (PB2, PB1, PA) aktiivisuus ja viruksen replikaatio lisääntyivät merkitsevästi. Tästä pääteltiin, että NF90 vaikuttaa negatiivisesti viruksen replikaatioon, siis hillitsee virusreplikaatiota.
Infektion alkuvaiheessa sekä viruksen NP että isäntäsolun NF90 lokalisoituvat yhdessä tumaan viruksella infektoituneissa soluissa , mikä viittaa siihen, että solun NF90 tekijän ja viruksen nukleoproteiinimonomeerien NP interaktio on varhainen tapahtuma viruksen replikaatiossa.
Kvantitatiivisella RT-PCR tutkimuksella voitiin osoittaa, että solun NF90 tekijä sääti alas SEKÄ viruksen replikaation ETTÄ mRNA transskription infektoituneissa soluissa.
Tulokset osoittavat, että NF90 estää influenssaviruksen replikaatiota infektion varhaisvaiheessa asettumalla suoraan interaktioon viruksen nukleoproteiinin (NP) kanssa
KOMMENTTINI:
Virukselle sen erittäin runsas NP-proteiinimonomeerijoukko toimii alustana, jonka varaan vRNA-käsittelytoiminnot replikaatiot, transkriptiot) rakentuvat ja jonka varaan uudet vRNA rihmat kiedotaan ja pakataan. NP on viruksen nukleokapsidin olennainen rakenne.
A-influenssaviruksen NP geeni on segmentti 5.
Nykyisessä A H1n1 sikainfluenssassa segmentti 5 on peräisin USA:ssa valliinneesta läntisestä sikainfluenssasta ja se pääsee helposti sekä transkriptioon ja replikaatioon. Muta tässä on tutkittu AH5N1 täyttä lintuvirusta, jonka keho ilmeiseti pystyy paremmin jarruttamaan. Ehkä jokin tällainen seikka on taustalla, että lintuvirus H5N1 ei pysty muuttumaan pandemiseksi nykyisillä lintugenomeillaan. Siis NP struktuuri voi olla ihmistumalle liian antigeeninen.
NP struktuurissa olevan aminohapon F- 412 tunnistaa solun puolusutusproteiini NF90 . Tunistaako se nykyisen H1N1 influenssaviruksen NP:n? Mikäli asia riippuu vain tuosta aminohaposta niin kyllä.
1..498
/organism="Influenza A virus (A/Moscow/02/2009(H1N1))"
/strain="A/Moscow/02/2009"
/serotype="H1N1"
/host="Homo sapiens"
/db_xref="taxon:650344"
/segment="5"
/country="Russia"
/collection_date="26-May-2009"
/note="lineage: swl"
Aminohapot 411- 420 ovat
tfsvqrnlpf
joten 412 on fenylalaniini (f) .
Influenssaviruksen geenit: PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS
Yksi seikka ontumatekijä NF90.
Mikä on ihmisen solun tumatekijä NF90?
Tumatekijä NF90 kuuluu tunnettuihin säätelijämolekyyleihin, jotka sitoutuvat antigeenivasteen tunnistuselementtiin ARRE-2 ja stimuloivat transkriptiota ja translaatiota normaalisolussa.
Mutta uutena tietona on, että tumatekijä NF90 voi myös negatiiviseti säädellä influenssaviruksen replikaatiota tekemällä vuorovaikutuksen influenssaviruksen nukleoproteiiniin NP, jos sitä esiintyy tumassa.
TUORE LÄHDE: http://www.citeulike.org/user/friendpine/article/4768934
Wang P, Song W et al. Nuclear factor 90, NF90, negatively regulates influenza virus replication by interacting with viral nucleoprotein. J Virol. 2009 Jun 3. [Epub ahead of print]
ISÄNTÄKEHON TEKIJÖIDEN ja VIRUSREPLIKAATION interaktiot ovat tärkeitä isäntäkehon vasteessa influenssavirukseen ja sen replikaation säätelyssä.
SOLUPROTEIINIA, jonka nimi on tumatekijä 90 (nuclear factor 90, NF90) , on löydetty ihmissoluista, joissa on esiintynyt ohimenevästi viruksen nukleoproteiinia NP ja tämä NF90 on löydetty samassa yhteydessä kun on puhdistettu viruksen H5N1 nukleoproteiinia NP käyttäen menetelmänä Mass Spectrometry (MALDI-TOF MS) analyysiä.
Koeputkessa NF90 ja NP-proteiinit saostuivat samanaikaisesti eräissä viljellyissä soluissa tai niitä esiintyi yksinään bakteerisoluissa tai kudosviljelmäsoluissa vastaavasti. Tämä löytö vahvisti, että NF90 ja NP välillä tapahtuu interaktiota itsenäisesti muista viruspartikkelin RNP proteiineista riippumatta.
Jos viruksen NP proteiinissa oli C-terminaalinen mutaatio (F412A) , interaktiota soluproteiiniin NF90 ei tapahtunut, joten tästä pääteltiin, että viruksen NP vaatii C- terminaalia sitoakseen NF90-proteiinia.
RNaasi V-käsittely ei asettanut esteitä NP ja NF90-proteiinien yhteiselle saostumiselle, mikä demonstroi, että niiden interaktio ei ollut riippuvainen RNA:sta.
Jos siRNA:lla tehdyn poistogeenisen NF90 vajeisen solun interaktioita tutkittiin, havaittiin, että siinä virusten polymeraasikompleksin (PB2, PB1, PA) aktiivisuus ja viruksen replikaatio lisääntyivät merkitsevästi. Tästä pääteltiin, että NF90 vaikuttaa negatiivisesti viruksen replikaatioon, siis hillitsee virusreplikaatiota.
Infektion alkuvaiheessa sekä viruksen NP että isäntäsolun NF90 lokalisoituvat yhdessä tumaan viruksella infektoituneissa soluissa , mikä viittaa siihen, että solun NF90 tekijän ja viruksen nukleoproteiinimonomeerien NP interaktio on varhainen tapahtuma viruksen replikaatiossa.
Kvantitatiivisella RT-PCR tutkimuksella voitiin osoittaa, että solun NF90 tekijä sääti alas SEKÄ viruksen replikaation ETTÄ mRNA transskription infektoituneissa soluissa.
Tulokset osoittavat, että NF90 estää influenssaviruksen replikaatiota infektion varhaisvaiheessa asettumalla suoraan interaktioon viruksen nukleoproteiinin (NP) kanssa
KOMMENTTINI:
Virukselle sen erittäin runsas NP-proteiinimonomeerijoukko toimii alustana, jonka varaan vRNA-käsittelytoiminnot replikaatiot, transkriptiot) rakentuvat ja jonka varaan uudet vRNA rihmat kiedotaan ja pakataan. NP on viruksen nukleokapsidin olennainen rakenne.
A-influenssaviruksen NP geeni on segmentti 5.
Nykyisessä A H1n1 sikainfluenssassa segmentti 5 on peräisin USA:ssa valliinneesta läntisestä sikainfluenssasta ja se pääsee helposti sekä transkriptioon ja replikaatioon. Muta tässä on tutkittu AH5N1 täyttä lintuvirusta, jonka keho ilmeiseti pystyy paremmin jarruttamaan. Ehkä jokin tällainen seikka on taustalla, että lintuvirus H5N1 ei pysty muuttumaan pandemiseksi nykyisillä lintugenomeillaan. Siis NP struktuuri voi olla ihmistumalle liian antigeeninen.
NP struktuurissa olevan aminohapon F- 412 tunnistaa solun puolusutusproteiini NF90 . Tunistaako se nykyisen H1N1 influenssaviruksen NP:n? Mikäli asia riippuu vain tuosta aminohaposta niin kyllä.
1..498
/organism="Influenza A virus (A/Moscow/02/2009(H1N1))"
/strain="A/Moscow/02/2009"
/serotype="H1N1"
/host="Homo sapiens"
/db_xref="taxon:650344"
/segment="5"
/country="Russia"
/collection_date="26-May-2009"
/note="lineage: swl"
Aminohapot 411- 420 ovat
tfsvqrnlpf
joten 412 on fenylalaniini (f) .
Influenssaviruksen geenit: PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS
A Influenssaviruksen kuva
A influenssan genomin replikaatiokompleksin osat ovat virionin sisällä nukleokapsidina (NC)
http://www.nature.com/scitable/nated/content/26874/10.1038_nrmicro1208-f1_mid_1.jpg
Genomi on segmentoitunut 8 eri osaan. Näistä jokainen osa (jokainen vRNA) voi olla eri lähteestä. Nykyinen H1N1 sikavirus omaa seuraavan sisäisen kalustuksen
1. segmentti . PB2 ( alunperin Pohjoisamerikkalaisesta lintuinfluenssasta käsin )
2. segmentti. PB1 (alunperin kausiluontoisesta ihmisen H3N2 influenssasta)
Tämä geeni voi lukeutua myös toiseksi proteiiniksi PB1-F2
3. segmentti. PA (alunperin Pohjois-amerikkalaisesta lintuinfluenssasta käsin)
4. segmentti. HA ( entisistä klassisista P- amer. sikainfluenssaviruksista käsin)
5. segmentti. NP (entisistä klassisista P-amer. sikainfluenssaviruksista käsin)
6. segmentti. NA (Euraasialaiseta sikainfluenssaviruksesta käsin)
7. segmentti. M (Euraasialaisesta sikainfluenssaviruksesta käsin). Tämä on kaksi-cistroninen ja tuotaa kaksi proteiinia M1 ja M2 .
8. segmentti. NS (entisista klassisista P-amer. sikainfluennsaviruksista käsin) . Tämä tuotaa kolme proteiinia NS1, NS2 ja NEP.
(KOMMENTTI: kuten näkyy, on hyvin epätoivottavaa, että tämä virus ottaisi lisäkalustusta lintuinfluenssasta päin, koska nykyisellään H1N1 ei ole vielä osoittanut kasvavaa letaliteettia, päinvastoin).
Influenssaviruksen geenit
PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS
http://www.nature.com/scitable/nated/content/26874/10.1038_nrmicro1208-f1_mid_1.jpg
{kind=link}
Genomi on segmentoitunut 8 eri osaan. Näistä jokainen osa (jokainen vRNA) voi olla eri lähteestä. Nykyinen H1N1 sikavirus omaa seuraavan sisäisen kalustuksen
1. segmentti . PB2 ( alunperin Pohjoisamerikkalaisesta lintuinfluenssasta käsin )
2. segmentti. PB1 (alunperin kausiluontoisesta ihmisen H3N2 influenssasta)
Tämä geeni voi lukeutua myös toiseksi proteiiniksi PB1-F2
3. segmentti. PA (alunperin Pohjois-amerikkalaisesta lintuinfluenssasta käsin)
4. segmentti. HA ( entisistä klassisista P- amer. sikainfluenssaviruksista käsin)
5. segmentti. NP (entisistä klassisista P-amer. sikainfluenssaviruksista käsin)
6. segmentti. NA (Euraasialaiseta sikainfluenssaviruksesta käsin)
7. segmentti. M (Euraasialaisesta sikainfluenssaviruksesta käsin). Tämä on kaksi-cistroninen ja tuotaa kaksi proteiinia M1 ja M2 .
8. segmentti. NS (entisista klassisista P-amer. sikainfluennsaviruksista käsin) . Tämä tuotaa kolme proteiinia NS1, NS2 ja NEP.
(KOMMENTTI: kuten näkyy, on hyvin epätoivottavaa, että tämä virus ottaisi lisäkalustusta lintuinfluenssasta päin, koska nykyisellään H1N1 ei ole vielä osoittanut kasvavaa letaliteettia, päinvastoin).
Influenssaviruksen geenit
PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS
lördag 13 juni 2009
M2-proteiinin mutaatio ja adamantaaniresistenssi
ADAMANTAANI-resistenssin arvio vuonna 2006 USA.ssa.
LÄHDE: CDC HEALTH ALERT 2006 CDC Recommends against the Use of Amantadine and Rimantadine for the Treatment or Prophylaxis of Influenza in the United States during the 2005–06 Influenza Season
Vuonna 2006 oli näyttöä kiertävien influenssakantojen amantadiini- ja rimantadiiniresistenssistä. Uutinen julkaistiin aikoinaan netissä seuraavassa lähteessä: Health Alert Network 2006 http://www.cdc.gov/flu/han011406.htm
PRIMÄÄRI strategia vastustaa vuosittaisen kausiluontoisen influenssan komplikaatioita, on vuosittainen rokotus
Taudin profylaksiassa ja influenssan hoidossa taas on sntiviruslääkeaineilla käyttöä.
ANTIVIRUSLÄÄKEAINEITA on kaksi luokkaa nykyisin
(1) M2-jonikanavan estäjät,
kuten kaksi adamantaania, amantadiini ja rimantadiini. (i.e., the two adamantanes amantadine and rimantadine) . Adamantaanit ovat tehokkaita vain influenssa A viruksia vastaan.
(2) Neuraminidaasin (NA) estäjät
kuten oseltamiviiri ja zanamaviiri (i.e., oseltamivir and zanamivir). Neuraminidaasin estäjät ovat tehokkaitta A ja B-influenssojen profylaksiassa ja hoidossa.
LÄÄKERESISTENSSIN muodostus
Vuosien 2005 ja 2006 influenssavirusten joukossa havaittiin USA:ssa resistenssiä adamantaaneille ja silloin tehtiin väliaikainen suositus, ettei näitä lääkkeitä käytettäisi sen aikaiseen kausi-influenssaan.
Amantadiinia käytetään myös Parkinosin taudin hoitoon ja siinä se on edelleen indisoitua käyttää.
(Kommentti: Olen tässä tosin sitä mieltä, että M2-kanavan aminohapporakenteessa on eräs neurotoksinen segmentti, jonka takia amantadiini voi olla profylaktinen, vaikka mitään erityistä M2-kanava vaikutusta muuten ei näkisi, niin ehkä viruksen aiheuttamat extrapyramidaaliset taudit jälkikomplikaatioina voitaisiin saada estymään. Tämä tuli mieleen kun katselin miltei kuin ”gliadiinia” näyttävää jaksoa M2 proteiinin C-terminaalisessa jaksossa. ).
Viruksen resistenssi adamantaaneille voi alkaa nopeasti hoidon aikana eräistä yksittäisten aminohappojen pistemutaatioista johtuen: M2-proteiinin aminohapot 26, 27, 30, 31, tai 34 voivat mutatoitua. Siitä voi seurata ristiresistenssiä sekä amantadiinia että rimantadiinia kohtaan. Mikään näistä aminohappomuutoksista ei heikennä adamantiiniresitenttien virusten tarttuvuutta eteenpäin.
Vuonna 2006 raportoitiin adamantiiniresistenttien virusten globaali prevalenssi. Se osoittautui nousseen merkitsevästi 1.9 %:sta 12.3%:iin 3 edeltäneen vuoden aikana.
USA:ssa lääkeresistenssi lisääntyi 1.9%:stä 14.5%:een vuosien 2004- 2005 influenssasesongin 6 ensimmäisen kuukauden aikana.
Vuonna 2005- 06 sesonkina otettiin 120 A influenssa H3N2 virusisolaattia 23 eri osavaltiosta CDC:n testeihin tammikuussa 2006. Näistä 106 ( 91%) käsitti M2-proteiinin rakenteessa aminohapon vaihtumisen asemassa 31 ja se vastasi amantadiini ja rimantadiiniresistenssistä. Kolme A (H1N1) kausiluontoista influenssavirusta testattiin myös ja niissä oli lääkeherkkyyttä.
Kaikki USA:ssa silloin testatut influenssavirukset osoittautuivat kyllä neuraminidaasi-inhibiittorille herkiksi. Tässä yhteydessä CDC esitti hälyttävän suosituksensa, ettei amantadiinia eikä rimantadiinia olisi käytetty koko influenssakautena 2005- 2006 USA:ssa, ei niin hoitoon eikä profylaksiaan. Mainittiin myös että suositus tultaisiin myöhemmin päivittämään tarvittaessa. Katsottiin, että vuosittainen rokote oli ensisijainen keino influenssaan liittyvän morbiditeetin ja mortaliteetin alentamiseen . Tästä annettiin sitten seuraavissa nettiosoitteissa jatkotiedot. www.cdc.gov/flu.
Neuraminidaasin estäjistä sai lisätietoa seuraavassa osoiteessa , joihin luvattiin päivitys-
www.cdc.gov/flu/protect/antiviral. These websites will be updated as new information becomes available.
13/06/2009 20:56
Influenssaviruksen geenit:
PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS
LÄHDE: CDC HEALTH ALERT 2006 CDC Recommends against the Use of Amantadine and Rimantadine for the Treatment or Prophylaxis of Influenza in the United States during the 2005–06 Influenza Season
Vuonna 2006 oli näyttöä kiertävien influenssakantojen amantadiini- ja rimantadiiniresistenssistä. Uutinen julkaistiin aikoinaan netissä seuraavassa lähteessä: Health Alert Network 2006 http://www.cdc.gov/flu/han011406.htm
PRIMÄÄRI strategia vastustaa vuosittaisen kausiluontoisen influenssan komplikaatioita, on vuosittainen rokotus
Taudin profylaksiassa ja influenssan hoidossa taas on sntiviruslääkeaineilla käyttöä.
ANTIVIRUSLÄÄKEAINEITA on kaksi luokkaa nykyisin
(1) M2-jonikanavan estäjät,
kuten kaksi adamantaania, amantadiini ja rimantadiini. (i.e., the two adamantanes amantadine and rimantadine) . Adamantaanit ovat tehokkaita vain influenssa A viruksia vastaan.
(2) Neuraminidaasin (NA) estäjät
kuten oseltamiviiri ja zanamaviiri (i.e., oseltamivir and zanamivir). Neuraminidaasin estäjät ovat tehokkaitta A ja B-influenssojen profylaksiassa ja hoidossa.
LÄÄKERESISTENSSIN muodostus
Vuosien 2005 ja 2006 influenssavirusten joukossa havaittiin USA:ssa resistenssiä adamantaaneille ja silloin tehtiin väliaikainen suositus, ettei näitä lääkkeitä käytettäisi sen aikaiseen kausi-influenssaan.
Amantadiinia käytetään myös Parkinosin taudin hoitoon ja siinä se on edelleen indisoitua käyttää.
(Kommentti: Olen tässä tosin sitä mieltä, että M2-kanavan aminohapporakenteessa on eräs neurotoksinen segmentti, jonka takia amantadiini voi olla profylaktinen, vaikka mitään erityistä M2-kanava vaikutusta muuten ei näkisi, niin ehkä viruksen aiheuttamat extrapyramidaaliset taudit jälkikomplikaatioina voitaisiin saada estymään. Tämä tuli mieleen kun katselin miltei kuin ”gliadiinia” näyttävää jaksoa M2 proteiinin C-terminaalisessa jaksossa. ).
Viruksen resistenssi adamantaaneille voi alkaa nopeasti hoidon aikana eräistä yksittäisten aminohappojen pistemutaatioista johtuen: M2-proteiinin aminohapot 26, 27, 30, 31, tai 34 voivat mutatoitua. Siitä voi seurata ristiresistenssiä sekä amantadiinia että rimantadiinia kohtaan. Mikään näistä aminohappomuutoksista ei heikennä adamantiiniresitenttien virusten tarttuvuutta eteenpäin.
Vuonna 2006 raportoitiin adamantiiniresistenttien virusten globaali prevalenssi. Se osoittautui nousseen merkitsevästi 1.9 %:sta 12.3%:iin 3 edeltäneen vuoden aikana.
USA:ssa lääkeresistenssi lisääntyi 1.9%:stä 14.5%:een vuosien 2004- 2005 influenssasesongin 6 ensimmäisen kuukauden aikana.
Vuonna 2005- 06 sesonkina otettiin 120 A influenssa H3N2 virusisolaattia 23 eri osavaltiosta CDC:n testeihin tammikuussa 2006. Näistä 106 ( 91%) käsitti M2-proteiinin rakenteessa aminohapon vaihtumisen asemassa 31 ja se vastasi amantadiini ja rimantadiiniresistenssistä. Kolme A (H1N1) kausiluontoista influenssavirusta testattiin myös ja niissä oli lääkeherkkyyttä.
Kaikki USA:ssa silloin testatut influenssavirukset osoittautuivat kyllä neuraminidaasi-inhibiittorille herkiksi. Tässä yhteydessä CDC esitti hälyttävän suosituksensa, ettei amantadiinia eikä rimantadiinia olisi käytetty koko influenssakautena 2005- 2006 USA:ssa, ei niin hoitoon eikä profylaksiaan. Mainittiin myös että suositus tultaisiin myöhemmin päivittämään tarvittaessa. Katsottiin, että vuosittainen rokote oli ensisijainen keino influenssaan liittyvän morbiditeetin ja mortaliteetin alentamiseen . Tästä annettiin sitten seuraavissa nettiosoitteissa jatkotiedot. www.cdc.gov/flu.
Neuraminidaasin estäjistä sai lisätietoa seuraavassa osoiteessa , joihin luvattiin päivitys-
www.cdc.gov/flu/protect/antiviral. These websites will be updated as new information becomes available.
13/06/2009 20:56
Influenssaviruksen geenit:
PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS
A H1N1 M2 proteiini ja amantadiini
VERTAILEN M2 proteiinia eri viruksilla kuten ihmisen pandeminen, A H1N1, sikainfluenssa H3N2 ja kanan A H5N1 ja Ihmiseen tarttunut A H5N1. Näyttää siltä että virukset jotka tulevat ihmispuolelle tulevat enemmän resistenteiksi amantadiinille asteittain. Mihin asia perustuu?
M2 proteiinin EKTODOMEENI on 24 aminohappoa, N-terminaali.
1-24
MSLLTEVETP
TRSEWECRCS
DSSD-
Transmembraanidomaani (TM) on 19 kpl aminohappoa
25-44
-PLVIAA
NIIGILHLIL
WITD-
Sytoplasminen C-terminaalinenpääty on 53 aminohappoa
45-97
-RLFFKC
IYRRFKYGLK
RGPSTEGVPE
SMREEYQQEQ
QSAVDVDDGH
FVNIELE"
Tämä 1- 97 sekvenssi M2 proteiinista on eräästä uudesta A H1N1 viruksesta ihmiseltä.
Amantadiinin ANTIVIRAALI VAIKUTUS kohdistuu vain neljään kohtaan M2 proteiinissa. . Amantadiinin ne vaikutukset, joille M2 voi olla altis, oletetaan tapahtuvan trans-Golgin alueella. Jos siinä kohtaa nimittäin pH ei pysyttele tarpeeksi matalana, muodostuva HA ei pääse muuttumaan funktiokelvolliseksi trimeeriksi, vaan aggrekoituu. Amantadiinin lisääminen infektion loppuvaiheeseen aiheuttaa sen takia prematuran konformationaalisen muutoksen HA rakenteessa. Trimeeri aggrekoituu , eikä kunnollisia virioneja muodostu.
Amantadiinin lisääminen infektion alkuvaiheessa vaikuttaa estäen M1 proteiinin dissosioitumista irti RNP. partikkelista, nukleokapsidista NC, jolloin M1-vRNP ei irtoa ja vRNP ei pääse ilmenemään tuman puolella. Jonikanavan tulisi nimittäin pitää yllä tiettyä edellytystä matalaan pH miljööseen ( protoneja H+), jossa miljöössä M1 dissosioituu RNP nukleokapsidista (NC) ja vapautuu siirtymään tumaan.
Mutta jos M2-proteiinin TM jaksossa on jokin mutaatio, amantadiinin vaikutukset estyvät.
M2-proteiini muodostuu siten, että se saa posttranslationaalisia muokkauksia, fosforylaatiota ja palmitylaatiota sytoplamiseen C-päätyyn, ja N-terminaalisiin cysteiineihin 17C ja 19C muodostuu disulfidisilta. Asylaatio ja disulfidisilta ei kuitenkaan vaikuta viruksen elinkykyyn eikä jonikanava-aktiviteettiin. Sanotaan, että TM- osa on alfa-helix ja siihen liittyy ne seikat, jotka muodostavat amantadiiniresistenssin. Amantadiinin sijoittumista alueelle Val -27 ja Ser-31 on raportoitu ja siinä se aiheuttaa steerisen blokin joninkanavan sisällä .
TM osan aminohapot ovat 20-44. (MUTTA Huomaa näissä H1N1 viruksissa on Seriini -31 mutatoitunut ja siinä on useimminkin N, asparagiini-31. Siis mutaatio S 31 N. Näyttää myös Valiinin Val (V) kohdalle tulevan jotain muuta joissain viruksissa, kuten isoleusiinia (V 27 I). Löysin konventionellin amantadiinille herkän jonikanavan erään kanan lintuinfluenssaviruksesta A H5N1. Siis mallia V-27 ja S-31.
Lisäksi Histidiini -37 (H-37) on virukselle olennainen jonikanavan funktiossa. Se näyttää olevan kaikissa viruksissa sama.
LÄHDE:
av Karl Maramorosch, Frederick A. Murphy, Aaron J … – 1999 – Medical – 487 sidor
ION CHANNEL PROTEIN The M2 protein is a 97-amino acid (aa) protein encoded by a … effect of amantadine and rimantadine on influenza virus infection
Influenssaviruksen M-geeni (M1/M2)
Influenssa A viruksen matrix proteiini M ja M geeni, segmentti 7.
M geeni on bicistroninen. Se tuottaa kahta proteiinia M1 ja M2. M1 on rakenneproteiini ja pääosa viruksen rakenteen suojaproteiineista. M2 on jonikanava .
1. MATRIXPROTEIINI M1 on transkription estäjä.
Matrixproteiini M1 tulee influenssaviruksen geenisegmentistä 7. Uusin tieto mitä siitä näkyy PubMed lähteessä on kaiketi eilispäivältä.
LÄHDE: Huang S, Chen J. Influenza A virus matrix protein 1 interacts with hTFIIIC102-s, a short isoform of the polypeptide 3 subunit of human general transcription factor IIIC. Arch Virol. 2009 Jun 12.
Influenssa A viruksen matrixproteiini 1 (M1) on multifunktionaalinen proteiini, jolla on virionin sisäseinämätehtävän lisäksi myös tärkeää osuutta viruksen replikaation (replication) onnistumisessa, uusien virionien oikeassa kokoontumisessa (assembly) ja vuorautumisessa ja sitten näiden virionien purkaantumisessa itsenäisiksi (budding)
http://www.microbiologybytes.com/virology/3035pics/Orthomyxo3.gif
http://www.schenectadyhistory.org/health/morris/1-100.jpg
Tämä tiedemiesryhmä halusi ottaa selvää, mitkä kaikki isäntäsolun sisällä esiintyvät proteiinit käyvät vuorovaikutukseen tämän virusproteiinin M1 kanssa, kun virus on purkanut sisältönsä isäntäsoluun. He tutkivat yhtätoista(11) itsenäistä proteiinia. Proteiinit olivat ihmissolun cDNA-kirjastosta seulottuja käyttämällä hiivan kahden hybridin tekniikkaa..
Näihin kuuluivat eräs transkriptiotekijä TF ( hTFIIIC102-s protein, encoding a short isoform of the TFIIIC102 subunit of the human TFIIIC transcription factor) Tiedemiehet tutkivat tarkasti mitten viruksen matrixproteiini M1 asettuu interaktioon ihmisen TFIIIC102-s proteiinin kanssa.
Interaktiossa tarvittiin seuraavat kohdat.
M1-proteiinista N-terminaalinen globulaarinen alue( aminohapot 1-164).
http://www2.ujf-grenoble.fr/pharmacie/laboratoires/gdrviro/grouperuigrok/archive/2001/figure2.gif
Ihmisen transkriptiotekijästä hTFIIIC102 viiden( tandem repeat, TR)tetra-tri-kopeptiditoiston (TPR1-5, aminohapot 149-362) alueTämä interaktio vahvistettiin kahdella eri menetelmällä. Lisäksi havaittiin, että HeLa-soluissa samanaikainen ilmenemä hTFIIIC102 transkriptiotekijää ja viruksen esti M1 –matrixproteiinia –translokoitumasta tumaan päin. M1 matrixproteiinia Yhteenvetona tutkijat sanovat, että ihmisen transkriptioproteiini TFIIIC102 voi vaikuttaa influenssaviruksen matrixproteiiniin M1 ja tästä saadaan uusia näkymiä M1-matrisproteiinin rooleihin influenssaviruksen ja isäntäsolun välisessä vuorovaikutuksessa.
2. MATRIX-proteiinista on paljon erikseen myös kirjoitettu, kuten tässä vuonna 2001 (Virus structure group)
http://www2.ujfgrenoble.fr/pharmacie/laboratoires/gdrviro/grouperuigrok/archive/2001/research.htm
LÄHDE: Florence Baudin, Isabelle Petit and rob Ruigrok in collaboration with Winfried Weissenhorn, Steffi Arzt (ESRF, Grenoble), and Wim Burmeister (UJF, Grenoble, visitor Cusack group).
Negatiivisen RNA-juosteen omaavien virusten matrixproteiini M ( tuhkarokon, ebolan, influenssan) omaa aivan välttämätöntä osaa siinä prosessissa, missä uusi virus partikkeli muodostuu infektoituneen solun pinnalla. Yksinkertaistettuna tämän ryhmän viruksilla on kuljetuskomponentit (TRANSPORT componentes) ja replikaatio (REPLICATION) -komponentit. Kuljetuspuolelle kuuluu glykoproteiinitapit joita viruksen lipidipinnalla on kalvorakenteeseen upotettuna( HA, NA) . Nämä proteiinit huolehtivat soluun sisään menosta ( ENTRY) , jotkut virukset myös tuhoavat solureseptoreita siten, että uudet vastamuodostuneet virionit voivat helpommin lähteä ulos solusta. Viruksen kalvo on fysikaalinen este, joka suojaa sisällä olevaa replikaatiokoneistoa ulkoisilta tekijöiltä.
http://s3.images.com/huge.55.278405.JPG
http://edoc.hu-berlin.de/dissertationen/rachakonda-p-sivaramakrishna-2005-02-15/HTML/image002.jpg
REPLIKAATIO-komponentti käsittää viruksen RNA nukleiinihapporakenteen, joka on sitoutuneena stokiometrisella tavalla nukleoproteiiniin NP. Spesifinen viruksen RNA:sta riippuva polymeraasikompleksi on siten sitoutuneena tähän N-RNA kompleksiin. (Sitä sanotaan polymeraasikompleksiksi ja influenssalla siihen kuuluu B2, PB1 ja PA) . Kaiken kaikkiaan totaalinen RNA ja proteiinikomponentit ovat yhteisnimeltään nukleokapsidi NC ja tämä rakenne vastaa viruksen vRNA:n replikaatiosta ja erilaisten mRNA-molekyylien tuotosta heti kun se on päässyt solun sisään ilman että mitään sen enempiä kuljetuskomponentteja ( TRANSPOR components) tarvittaisiin.
VIRUSPARTIKKELIN sisällä matrixproteiinit(M) liimaavat TRANSPORT- ja REPLIKAATIO- komponentteja yhteen . Jotta M pystyisi tekemään sillä tavalla, sen täytyy tehdä kontakti sekä glykoproteiinien sytoplasmisiin päätyihin lipidikalvossa, jolla on spesifinen rakenne (raft) että nukleokapsidin kanssa ja lisäski sisäisesti, sillä se on muodoltaan polymeerinen matrixprotiini, joka vaikuttaa virionin muotoon.
Tämä työryhmä on tutkinut influenssaproteiineja 10 vuotta. ja he ovat myös vertailleet erään toisen työryhmän tietoihin Ebola-viruksen matrixproteiinista (Winfried Weissenhorn)
Influenssaviruksen M-proteiinin spontaaniproteolyysissä havaitaan N-terminaalinen domaani, jossa on aminohappoja 1- 164. Tämä julkaistiin vuonna 2000. Bakteerissa tuotettiin kokomittainen M1 proteiini. Sen N-terminaali domaani ja C-terminaalinen domaani(165-252). Havaittiin, että vain koko pitkä M1 ja sen N-terminaalinen domaani sitoutuivat negatiivisesti varautuneihin liposomeihin ( mikä on yksinkertainen oikean viruksen kalvon vastine kokeissa) ja että vain kokopitkä M1 ja sen C-terminaalinen osa sitoutuivat viraalisen RNP sauvaan.
Kuitenkin vain kokopitkällä M1-molekyylillä oli aktiivisuutta, jolla se esti transkriptiota.
Influenssaviruksen geenistö:
PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS
Toisistaan riippumattomissa kokeissa selviteltiin edelleen asiaa. Koeputkioloissa transkriptio estyi nonspesifisellä M1 proteiinin polymerisaatiolla/aggrekaatiolla RNP-partikkeliin TAI M1 proteiinilla, joka sitoutui toiseen M1-proteiiniin RNP.ssä pikemminkin kuin sitoutumalla NP proteiinin johonkin spesifiseen aktiiviin kohtaan tai polymeraasiin.
Tämä viitaisi siihen, että tämä transkriptiota estävä aktiivisuus, jota kauan jo ollaan kuvattu matrixproteiinin eräänä spesifisena aktiivisuutena, ei näytä olevan spesifinen ollenkaan vaan sivuvaikutus proteiinin luonnollsesta taipumuksesta sitoutua nukleokapsidiin NC ja itse polymerisoitua.
Sen mukaan mitä voimme tästä itse päätellä M-proteiini on aina isoloituna itse viruksesta.
Entä EBOLAN matrixproteiini? VP40, se tuottuu yhtenä domaanina, liukoisena proteiinina, joka destabilisoituessaan tai joutuessaan kontaktiin negatiivissti varautuneiten kalvojen kanssa, avautuu muodostaen kaksi domaania. Sen muutoksen jälkeen VP40 muodostaa oligomeeririnkuloita ja pstyy sitoutumaan kalvoihin. Ei ole vielä testattu pystyykö se myös kiinnittymään nukleokapsideihin. VP40:n kalvoihin sitoutuva aktiivisuus pysyttelee sen C-terminaalisessa domaanissa , kun taas influenssaviruksen matrix-proteiinin M1 vastaava aktiivisuus on N-terminaalisessa domaanissa ja nukleokapsidinkin suhteen käyttäytyy eri tavalla. Vaikuttaa siltä, että näillä kahdella rakenteella on todellakin samoja tehtäviä, mutta ne eivät ole millään muotoa homologisia, mikä näkyy jo niiden atomsesta rakenteesta.
3. VIRUKSEN M-geeni, 8 segmentin geeni tuottaa proteiinit M1 ja M2
M-proteiinien M1 rakenteet ovat virionin päämateriaalia,suojaavaa matrixproteiinia. M2 taas on integralinen membraaniproteiini, jonka tehtvä on tietyssä vaiheessa toimia jonikanavana.
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=249043
Kun on analysoitu M1 ja M2 geenejä ja niiden geenituotteita proteiineja, on havaittu niillä olevan hyvin erilaiset evoluutiotavat. Jos taas verrataan muiden influenssageenien evoluutiotapoihin (esim PB2 ja NP geeneihin), niin M1 ja M2 geenit evoloituvat suhteellisen hitaasti, erityisesti M1 geeni on hidas evoluutiossaan. M1 ja M2 geenituotteet koodautuvat eri lukuraameista (reading frames) mutta toisiaan kattavista (overlapping reading frames) kohdista. Kun näitä tutkittiin, havaittiin, että M1 proteiinit evoloituvat hyvin hitaasti kaikissa linjoissa, kun taas M2 proteiinit osoittavat merkitsevää evoluutiota ihmis-ja sikainfluenssalinjoissa, mutta käytännöllisesti katsoen ei mitään evoluutiota lintuinfluenssalinjoissa.
Evoluutiotahti M1 proteiineissa oli paljon matalampi kuin M2 proteiineissa ja muissa influenssan sisäosan proteiineissa (PB2 ja NP). Samalla M2 proteiineillä taas muihin pintaproteiinehin (esim. H3HA) verrattuna ei ollut niin nopea evoluutio.
Tutkijoiden tulokset osoittavat , että A-influenssaviruksilla bi-cistronisen geenin yhden proteiinin evoluutiolla on merkitystä sen muiden proteiinien evoluutioon
13/06/2009
12:15
M geeni on bicistroninen. Se tuottaa kahta proteiinia M1 ja M2. M1 on rakenneproteiini ja pääosa viruksen rakenteen suojaproteiineista. M2 on jonikanava .
1. MATRIXPROTEIINI M1 on transkription estäjä.
Matrixproteiini M1 tulee influenssaviruksen geenisegmentistä 7. Uusin tieto mitä siitä näkyy PubMed lähteessä on kaiketi eilispäivältä.
LÄHDE: Huang S, Chen J. Influenza A virus matrix protein 1 interacts with hTFIIIC102-s, a short isoform of the polypeptide 3 subunit of human general transcription factor IIIC. Arch Virol. 2009 Jun 12.
Influenssa A viruksen matrixproteiini 1 (M1) on multifunktionaalinen proteiini, jolla on virionin sisäseinämätehtävän lisäksi myös tärkeää osuutta viruksen replikaation (replication) onnistumisessa, uusien virionien oikeassa kokoontumisessa (assembly) ja vuorautumisessa ja sitten näiden virionien purkaantumisessa itsenäisiksi (budding)
http://www.microbiologybytes.com/virology/3035pics/Orthomyxo3.gif
{kind=link}
http://www.schenectadyhistory.org/health/morris/1-100.jpg
{kind=link}
Tämä tiedemiesryhmä halusi ottaa selvää, mitkä kaikki isäntäsolun sisällä esiintyvät proteiinit käyvät vuorovaikutukseen tämän virusproteiinin M1 kanssa, kun virus on purkanut sisältönsä isäntäsoluun. He tutkivat yhtätoista(11) itsenäistä proteiinia. Proteiinit olivat ihmissolun cDNA-kirjastosta seulottuja käyttämällä hiivan kahden hybridin tekniikkaa..
Näihin kuuluivat eräs transkriptiotekijä TF ( hTFIIIC102-s protein, encoding a short isoform of the TFIIIC102 subunit of the human TFIIIC transcription factor) Tiedemiehet tutkivat tarkasti mitten viruksen matrixproteiini M1 asettuu interaktioon ihmisen TFIIIC102-s proteiinin kanssa.
Interaktiossa tarvittiin seuraavat kohdat.
M1-proteiinista N-terminaalinen globulaarinen alue( aminohapot 1-164).
http://www2.ujf-grenoble.fr/pharmacie/laboratoires/gdrviro/grouperuigrok/archive/2001/figure2.gif
Ihmisen transkriptiotekijästä hTFIIIC102 viiden( tandem repeat, TR)tetra-tri-kopeptiditoiston (TPR1-5, aminohapot 149-362) alueTämä interaktio vahvistettiin kahdella eri menetelmällä. Lisäksi havaittiin, että HeLa-soluissa samanaikainen ilmenemä hTFIIIC102 transkriptiotekijää ja viruksen esti M1 –matrixproteiinia –translokoitumasta tumaan päin. M1 matrixproteiinia Yhteenvetona tutkijat sanovat, että ihmisen transkriptioproteiini TFIIIC102 voi vaikuttaa influenssaviruksen matrixproteiiniin M1 ja tästä saadaan uusia näkymiä M1-matrisproteiinin rooleihin influenssaviruksen ja isäntäsolun välisessä vuorovaikutuksessa.
2. MATRIX-proteiinista on paljon erikseen myös kirjoitettu, kuten tässä vuonna 2001 (Virus structure group)
http://www2.ujfgrenoble.fr/pharmacie/laboratoires/gdrviro/grouperuigrok/archive/2001/research.htm
LÄHDE: Florence Baudin, Isabelle Petit and rob Ruigrok in collaboration with Winfried Weissenhorn, Steffi Arzt (ESRF, Grenoble), and Wim Burmeister (UJF, Grenoble, visitor Cusack group).
Negatiivisen RNA-juosteen omaavien virusten matrixproteiini M ( tuhkarokon, ebolan, influenssan) omaa aivan välttämätöntä osaa siinä prosessissa, missä uusi virus partikkeli muodostuu infektoituneen solun pinnalla. Yksinkertaistettuna tämän ryhmän viruksilla on kuljetuskomponentit (TRANSPORT componentes) ja replikaatio (REPLICATION) -komponentit. Kuljetuspuolelle kuuluu glykoproteiinitapit joita viruksen lipidipinnalla on kalvorakenteeseen upotettuna( HA, NA) . Nämä proteiinit huolehtivat soluun sisään menosta ( ENTRY) , jotkut virukset myös tuhoavat solureseptoreita siten, että uudet vastamuodostuneet virionit voivat helpommin lähteä ulos solusta. Viruksen kalvo on fysikaalinen este, joka suojaa sisällä olevaa replikaatiokoneistoa ulkoisilta tekijöiltä.
http://s3.images.com/huge.55.278405.JPG
{kind=link}
http://edoc.hu-berlin.de/dissertationen/rachakonda-p-sivaramakrishna-2005-02-15/HTML/image002.jpg
REPLIKAATIO-komponentti käsittää viruksen RNA nukleiinihapporakenteen, joka on sitoutuneena stokiometrisella tavalla nukleoproteiiniin NP. Spesifinen viruksen RNA:sta riippuva polymeraasikompleksi on siten sitoutuneena tähän N-RNA kompleksiin. (Sitä sanotaan polymeraasikompleksiksi ja influenssalla siihen kuuluu B2, PB1 ja PA) . Kaiken kaikkiaan totaalinen RNA ja proteiinikomponentit ovat yhteisnimeltään nukleokapsidi NC ja tämä rakenne vastaa viruksen vRNA:n replikaatiosta ja erilaisten mRNA-molekyylien tuotosta heti kun se on päässyt solun sisään ilman että mitään sen enempiä kuljetuskomponentteja ( TRANSPOR components) tarvittaisiin.
VIRUSPARTIKKELIN sisällä matrixproteiinit(M) liimaavat TRANSPORT- ja REPLIKAATIO- komponentteja yhteen . Jotta M pystyisi tekemään sillä tavalla, sen täytyy tehdä kontakti sekä glykoproteiinien sytoplasmisiin päätyihin lipidikalvossa, jolla on spesifinen rakenne (raft) että nukleokapsidin kanssa ja lisäski sisäisesti, sillä se on muodoltaan polymeerinen matrixprotiini, joka vaikuttaa virionin muotoon.
Tämä työryhmä on tutkinut influenssaproteiineja 10 vuotta. ja he ovat myös vertailleet erään toisen työryhmän tietoihin Ebola-viruksen matrixproteiinista (Winfried Weissenhorn)
Influenssaviruksen M-proteiinin spontaaniproteolyysissä havaitaan N-terminaalinen domaani, jossa on aminohappoja 1- 164. Tämä julkaistiin vuonna 2000. Bakteerissa tuotettiin kokomittainen M1 proteiini. Sen N-terminaali domaani ja C-terminaalinen domaani(165-252). Havaittiin, että vain koko pitkä M1 ja sen N-terminaalinen domaani sitoutuivat negatiivisesti varautuneihin liposomeihin ( mikä on yksinkertainen oikean viruksen kalvon vastine kokeissa) ja että vain kokopitkä M1 ja sen C-terminaalinen osa sitoutuivat viraalisen RNP sauvaan.
Kuitenkin vain kokopitkällä M1-molekyylillä oli aktiivisuutta, jolla se esti transkriptiota.
Influenssaviruksen geenistö:
PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS
Toisistaan riippumattomissa kokeissa selviteltiin edelleen asiaa. Koeputkioloissa transkriptio estyi nonspesifisellä M1 proteiinin polymerisaatiolla/aggrekaatiolla RNP-partikkeliin TAI M1 proteiinilla, joka sitoutui toiseen M1-proteiiniin RNP.ssä pikemminkin kuin sitoutumalla NP proteiinin johonkin spesifiseen aktiiviin kohtaan tai polymeraasiin.
Tämä viitaisi siihen, että tämä transkriptiota estävä aktiivisuus, jota kauan jo ollaan kuvattu matrixproteiinin eräänä spesifisena aktiivisuutena, ei näytä olevan spesifinen ollenkaan vaan sivuvaikutus proteiinin luonnollsesta taipumuksesta sitoutua nukleokapsidiin NC ja itse polymerisoitua.
Sen mukaan mitä voimme tästä itse päätellä M-proteiini on aina isoloituna itse viruksesta.
Entä EBOLAN matrixproteiini? VP40, se tuottuu yhtenä domaanina, liukoisena proteiinina, joka destabilisoituessaan tai joutuessaan kontaktiin negatiivissti varautuneiten kalvojen kanssa, avautuu muodostaen kaksi domaania. Sen muutoksen jälkeen VP40 muodostaa oligomeeririnkuloita ja pstyy sitoutumaan kalvoihin. Ei ole vielä testattu pystyykö se myös kiinnittymään nukleokapsideihin. VP40:n kalvoihin sitoutuva aktiivisuus pysyttelee sen C-terminaalisessa domaanissa , kun taas influenssaviruksen matrix-proteiinin M1 vastaava aktiivisuus on N-terminaalisessa domaanissa ja nukleokapsidinkin suhteen käyttäytyy eri tavalla. Vaikuttaa siltä, että näillä kahdella rakenteella on todellakin samoja tehtäviä, mutta ne eivät ole millään muotoa homologisia, mikä näkyy jo niiden atomsesta rakenteesta.
3. VIRUKSEN M-geeni, 8 segmentin geeni tuottaa proteiinit M1 ja M2
M-proteiinien M1 rakenteet ovat virionin päämateriaalia,suojaavaa matrixproteiinia. M2 taas on integralinen membraaniproteiini, jonka tehtvä on tietyssä vaiheessa toimia jonikanavana.
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=249043
Kun on analysoitu M1 ja M2 geenejä ja niiden geenituotteita proteiineja, on havaittu niillä olevan hyvin erilaiset evoluutiotavat. Jos taas verrataan muiden influenssageenien evoluutiotapoihin (esim PB2 ja NP geeneihin), niin M1 ja M2 geenit evoloituvat suhteellisen hitaasti, erityisesti M1 geeni on hidas evoluutiossaan. M1 ja M2 geenituotteet koodautuvat eri lukuraameista (reading frames) mutta toisiaan kattavista (overlapping reading frames) kohdista. Kun näitä tutkittiin, havaittiin, että M1 proteiinit evoloituvat hyvin hitaasti kaikissa linjoissa, kun taas M2 proteiinit osoittavat merkitsevää evoluutiota ihmis-ja sikainfluenssalinjoissa, mutta käytännöllisesti katsoen ei mitään evoluutiota lintuinfluenssalinjoissa.
Evoluutiotahti M1 proteiineissa oli paljon matalampi kuin M2 proteiineissa ja muissa influenssan sisäosan proteiineissa (PB2 ja NP). Samalla M2 proteiineillä taas muihin pintaproteiinehin (esim. H3HA) verrattuna ei ollut niin nopea evoluutio.
Tutkijoiden tulokset osoittavat , että A-influenssaviruksilla bi-cistronisen geenin yhden proteiinin evoluutiolla on merkitystä sen muiden proteiinien evoluutioon
13/06/2009
12:15
Influenssaviruksen dynaaminen genomi
Influenssaviruksen genomi on evoluutioltaan dynaaminen
Influenssavirus sopeutuu (adaptoituu) huomattavan hyvin ympäristöihinsä ja sen takia pysyy pitkät ajat ihmisväestöissä. Ihmisinfluenssa (human) A virus pystyy viihtymään vielä silloinkin ihmisväestössä, kun jo on rokotteita laajaltikin saatavilla ja rokotteista huolimatta se aiheuttaa runsaasti morbiditeettia, sairautta ja mortaliteettia, kuolemaa. Tämä virustyyppi mutatoituu vuodesta toiseen ja olemassaolevat rokotteet tulevat pian tehottomiksi, minkä takia pitää uuteen rokotteeseen valita taas uusia kantoja.
Isoja antigeenin vaihtumisia ( antigenic shift) voi joskus myös tapahtua ja tällöin ihmiskunnalla on huono protektiivinen immuniteetti ja suojaamaton väestö voi silloin joutua laajemman globaalin epidemian kouriin. Näitä ovat pandemiat. Pahin pandemia, mitä vuosisadan sisällä on ollut, oli vuoden 1918 Espanjan tauti (H1N1) ja se niitti 30-50 miljoonaa ihmistä. Sitten muita pahoja pandemioita oli vuoden 1957 aasialainen influenssa (H2N2) ja vuoden 1968 hongkongilainen influenssa (H3N2).
Koska influenssaviruksen genomi on niin suuresti muuntelevainen, tiedemiehet ovat alkaneet pitää sen rakenteesta hyvin tarkkaa selkoa. Tässä allamainitussa tutkimuksessa työryhmä selvitti 209 ihmisen A-influenssaviruksen genomisen rakenteen täydellisesti. He selvittivät kaiken kaikkiaan 2 821 103 nukleotidiä (nt). Täten he pystyivät lisäämään tiedemiesten luetteloihin ison joukon uusia selviteltyjä A-influenssavirusgenomeja ja lisäksi he totesivat näissä 209 A-influenssaviruksessa useita anomalioita, poikkeavuuksia, mikä viittaa siihen, että influenssan tarttuminen ja evoluutio on luonteeltaan dynaamista.Tähän evoluutioon tutkimus toi uutta valoa, samoin viruksen tarttumismalleihin ihmis- ja eläinkunanssa. Tutkijat antoivat saamansa tiedot viiveettä yleisiin tietueisiin.
LÄHDE: Ghedin E et al. Large-scale sequencing of human influenza reveals the dynamic nature of viral genome evolution.. Nature. 2005 Oct 5;
TIETOJA INFLUENSSAVIRUKSEN GENOMISTA
http://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=11320
http://www.ncbi.nlm.nih.gov/About/disclaimer.html
Kaikki 11 virus proteiinia koodittuvat 8 geenillä, jotka ovat RNA segmenttejä.
PB2,
PB1, PB1-F2,
PA,
HA,
NP,
NA,
M1, M2,
NS1 ja NS2.
http://www.cdc.gov/ncidod/eid/vol12no09/06-0276.htm#afigure1
Sitaatti, suomennos:
Tutkijat olivat katsoneet yllämainitussa lähteessä 306 ihmis ja 95 lintuinfluenssagenomia, mikä osoitti että 228 aminohappoa 4591 aminohaposta osoittivat erilaisuuksia näiden eri lajin virusten kesken. Sitten he tutkivat 15 785 proteiinisekvenssiä määrittääkseen näiden signatuurien yleisyyden ja löysivät 52 lajista riippuva asemaa.
Spesifiset mutaatiot lintuviruksen näissä kohdissa saivat lintuviruksen hankkimaan kykyä muuttua ihmisvirukseksi.
Moni signatuuri löytyi NP, PA ja PB2 geeneissä( viruksen ribonukleoproteiinissa vRNP) ja ovat enimmäkseen paikallistuneina funktionaalisiin domaaneihin, jotka toimivat RNP-RNP interaktioissa, joilla on merkitystä viruksen replikaatiolle.
Kun tutkijat tarkastelivat 21 ihmisistä eristettyä lintuinfluenssavirusgenomia, he havaitsivat, että 19 viruksella ilmeni yksi tai useampi lajiin assosioituva muutos. Seitsemällä niistä oli 2 tai useampi substituutio. Histogrammit osoittivat, että parien sekventoivassa vertailussa NP oli kaikkein useimmin epäyhtenäinen( disjointed) ihmis- ja lintuviruksen kesken , sen jälkeen PA ja sitten PB2.
KOMMENTTI: Nykyisellä pandemisella "sekaviruksella"vRNP rakenteesta
NP on länsimaisista sikainfluenssalajeista.
PA on lintuinfluenssasta.
PB2 on lintuinfluenssasta.
(vRNP-rakenteeseen kuuluva PB1 on ihmisviruksesta) .
Influenssavirus sopeutuu (adaptoituu) huomattavan hyvin ympäristöihinsä ja sen takia pysyy pitkät ajat ihmisväestöissä. Ihmisinfluenssa (human) A virus pystyy viihtymään vielä silloinkin ihmisväestössä, kun jo on rokotteita laajaltikin saatavilla ja rokotteista huolimatta se aiheuttaa runsaasti morbiditeettia, sairautta ja mortaliteettia, kuolemaa. Tämä virustyyppi mutatoituu vuodesta toiseen ja olemassaolevat rokotteet tulevat pian tehottomiksi, minkä takia pitää uuteen rokotteeseen valita taas uusia kantoja.
Isoja antigeenin vaihtumisia ( antigenic shift) voi joskus myös tapahtua ja tällöin ihmiskunnalla on huono protektiivinen immuniteetti ja suojaamaton väestö voi silloin joutua laajemman globaalin epidemian kouriin. Näitä ovat pandemiat. Pahin pandemia, mitä vuosisadan sisällä on ollut, oli vuoden 1918 Espanjan tauti (H1N1) ja se niitti 30-50 miljoonaa ihmistä. Sitten muita pahoja pandemioita oli vuoden 1957 aasialainen influenssa (H2N2) ja vuoden 1968 hongkongilainen influenssa (H3N2).
Koska influenssaviruksen genomi on niin suuresti muuntelevainen, tiedemiehet ovat alkaneet pitää sen rakenteesta hyvin tarkkaa selkoa. Tässä allamainitussa tutkimuksessa työryhmä selvitti 209 ihmisen A-influenssaviruksen genomisen rakenteen täydellisesti. He selvittivät kaiken kaikkiaan 2 821 103 nukleotidiä (nt). Täten he pystyivät lisäämään tiedemiesten luetteloihin ison joukon uusia selviteltyjä A-influenssavirusgenomeja ja lisäksi he totesivat näissä 209 A-influenssaviruksessa useita anomalioita, poikkeavuuksia, mikä viittaa siihen, että influenssan tarttuminen ja evoluutio on luonteeltaan dynaamista.Tähän evoluutioon tutkimus toi uutta valoa, samoin viruksen tarttumismalleihin ihmis- ja eläinkunanssa. Tutkijat antoivat saamansa tiedot viiveettä yleisiin tietueisiin.
LÄHDE: Ghedin E et al. Large-scale sequencing of human influenza reveals the dynamic nature of viral genome evolution.. Nature. 2005 Oct 5;
TIETOJA INFLUENSSAVIRUKSEN GENOMISTA
http://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=11320
http://www.ncbi.nlm.nih.gov/About/disclaimer.html
Kaikki 11 virus proteiinia koodittuvat 8 geenillä, jotka ovat RNA segmenttejä.
PB2,
PB1, PB1-F2,
PA,
HA,
NP,
NA,
M1, M2,
NS1 ja NS2.
http://www.cdc.gov/ncidod/eid/vol12no09/06-0276.htm#afigure1
Sitaatti, suomennos:
Tutkijat olivat katsoneet yllämainitussa lähteessä 306 ihmis ja 95 lintuinfluenssagenomia, mikä osoitti että 228 aminohappoa 4591 aminohaposta osoittivat erilaisuuksia näiden eri lajin virusten kesken. Sitten he tutkivat 15 785 proteiinisekvenssiä määrittääkseen näiden signatuurien yleisyyden ja löysivät 52 lajista riippuva asemaa.
Spesifiset mutaatiot lintuviruksen näissä kohdissa saivat lintuviruksen hankkimaan kykyä muuttua ihmisvirukseksi.
Moni signatuuri löytyi NP, PA ja PB2 geeneissä( viruksen ribonukleoproteiinissa vRNP) ja ovat enimmäkseen paikallistuneina funktionaalisiin domaaneihin, jotka toimivat RNP-RNP interaktioissa, joilla on merkitystä viruksen replikaatiolle.
Kun tutkijat tarkastelivat 21 ihmisistä eristettyä lintuinfluenssavirusgenomia, he havaitsivat, että 19 viruksella ilmeni yksi tai useampi lajiin assosioituva muutos. Seitsemällä niistä oli 2 tai useampi substituutio. Histogrammit osoittivat, että parien sekventoivassa vertailussa NP oli kaikkein useimmin epäyhtenäinen( disjointed) ihmis- ja lintuviruksen kesken , sen jälkeen PA ja sitten PB2.
KOMMENTTI: Nykyisellä pandemisella "sekaviruksella"vRNP rakenteesta
NP on länsimaisista sikainfluenssalajeista.
PA on lintuinfluenssasta.
PB2 on lintuinfluenssasta.
(vRNP-rakenteeseen kuuluva PB1 on ihmisviruksesta) .
Pidä huolta keuhkoistasi ja yleiskunnostasi
Oseltamivir ( Tamiflu) tehostaa omien anti-influenssa-aktiviteettia omaavien luonnollisten immuuniproteiinien toimintaa, kuten keuhkojen omien surfaktanttien, pintajännitystä alentavien keuhkosolutuotteiden toimintaa,
KEUHKOSURFAKTANTTIEN OSUUS
Influenssa A virusta (IAV) luonnostaan estäviä tekijöitä ovat surfaktanttikollektiinit A ja D ( SP-A ja SP-D) sekä glykoproteiini 340 (gp340), jossa on paljon scavencer- reseptoreita ( vapaitten radikaalien hävittäjien reseptoreita).
http://www.nature.com/nri/journal/v5/n1/images/nri1528-f1.jpg
http://www.nature.com/nri/journal/v5/n1/fig_tab/nri1528_F1.html
Näillä on seuraavanlaiset tehot: Tehokkain on: SP-D, sitten gp340 ja sitten SP-A.
SP-D sitoutuu kalsiumista riippuvalla tavalla viruksen hemagglutiniinin (HA) tai viruksen neuraminidaasin (NA) hiilihydraattirakenteisiin.
Päinvastoin kuin SP-D toimivat gp340 ja SP-A ja samalla tavalla kuten musiinit siinä mielessä, että ne tarjoavat sialiinihappoligandeja, jotka kiinnittyvät viruksen hemagglutiniiniin (HA).
Tässä tutkimuksessa nämä SP-D, SP-A ja gp340 osoittivat ko-operaatiota antiviraalisissa interaktioissaan. Yhteistoimintavaikutukset oli parhainta viruksen aggregoitumista vastaan., mutta ilmenivät myös jonkin verran joinain havaittuna hemagglutinaation inhibitioina ja viruksen neutralisoimisina.
Monoklonaalisilla vasta-aineilla (MoAB) tutkittiin edelleen gp340 ja SP-D-surfaktatanttiproteiinin välistä sitoutumista. Vaikka gp340 pystyi sitoutumaan SPD:hen muussakin kuin mannaania sitovassa kohdassa, ei tämä gp340 ja SP-D-sitoutuminen tuottanut kuitenkaan mitään lisäetua antivirusinteraktioihin.
SP-D ja musiini ( mucus) osoittivat interaktiivista yhteistoimintaa ilmeisesti sen takia, että SP-D esti viruksen neuraminidaasia (NA).
Markkinoilla oleva NA-inhibiittori oseltamivir ( Tamiflu) osoitti samankaltaista vaikutusta ja lisäsi myös SP-A surfaktantin (keuhkojen pintajännitystä alentavan tekijän) neutraloivaa toimintaa ja bronkoalveolaarista huuhtelunestettä.
Täten oseltamivir on yhteistyössä luonnollisten immuuniproteiinien kanssa estäessään epiteelisolujen initiaalista infektiota.
LÄHDE:
White MR, Crouch E et.al. Cooperative anti-influenza activities of respiratory innate immune proteins and neuraminidase inhibitor. Am J Physiol Lung Cell Mol Physiol. 2005 May;288(5):L831-40. Epub 2004 Dec 17.
Department of Medicine, Section of Hematology/Oncology, Boston University, School of Medicine, Boston, MA
Terminologia:
surfactant collectins, pintajännitystä alentavat kollektiinit
surfactant proteins A and D (SP-A and D), surfaktanttiproteiinit SP-A ja SP-D
http://ising.phys.cwru.edu/surfactants/proteins.html
scavenger receptor-rich glycoprotein 340 (gp340). glykoproteiini 340, jossa on vapaita radikaaleja pyydystäviä reseptoreita paljon
mucins, musiinit, limaaineet
sialic acid ligands, sialihappoon asettuvia molekyylejä
oseltamivir, neuraminidaseinhibitor, neuraminidaasin estäjä
PIDÄ HUOLTA OMISTA LUONNOLLISISTA SURFAKTANTEISTASI. Tamiflu ja Relenza auttavat ehkä siksi, että lääke tekee yhteistyötä oman puolustuksen keuhkosurfaktanttien ja keuhkomusiinien kanssa ja niiden saaatua apua osetamiviriltä keuhkojen toipuminen influenssasta on parempi.
SURFAKTANTTIEN muodostuminen on parasta, kun on sopivasti ruokaa ja juomaa ja ehkä erityisesti maitotuotteita, kermaa, maitorasvoja, kasvisrasvoja, sipulia, C-vitamiinia, acetylcysteiiniä ( poretablettina käytettävä lääke) . Hyvällä yleiskunnolla ja sopivalla ravinnolla ja antioksidanteilla voi tehostaa luonnollisen puolustuksensa surfaktanttien muodostusta.
On eduksi, jos ei polta tupakkaa.
Influenssan ja kuivan yskän tullen on eduksi käyttää jotain yskänärsytystä hillitsevää lääkettä.
KEUHKOSURFAKTANTTIEN OSUUS
Influenssa A virusta (IAV) luonnostaan estäviä tekijöitä ovat surfaktanttikollektiinit A ja D ( SP-A ja SP-D) sekä glykoproteiini 340 (gp340), jossa on paljon scavencer- reseptoreita ( vapaitten radikaalien hävittäjien reseptoreita).
http://www.nature.com/nri/journal/v5/n1/images/nri1528-f1.jpg
{kind=link}
http://www.nature.com/nri/journal/v5/n1/fig_tab/nri1528_F1.html
Näillä on seuraavanlaiset tehot: Tehokkain on: SP-D, sitten gp340 ja sitten SP-A.
SP-D sitoutuu kalsiumista riippuvalla tavalla viruksen hemagglutiniinin (HA) tai viruksen neuraminidaasin (NA) hiilihydraattirakenteisiin.
Päinvastoin kuin SP-D toimivat gp340 ja SP-A ja samalla tavalla kuten musiinit siinä mielessä, että ne tarjoavat sialiinihappoligandeja, jotka kiinnittyvät viruksen hemagglutiniiniin (HA).
Tässä tutkimuksessa nämä SP-D, SP-A ja gp340 osoittivat ko-operaatiota antiviraalisissa interaktioissaan. Yhteistoimintavaikutukset oli parhainta viruksen aggregoitumista vastaan., mutta ilmenivät myös jonkin verran joinain havaittuna hemagglutinaation inhibitioina ja viruksen neutralisoimisina.
Monoklonaalisilla vasta-aineilla (MoAB) tutkittiin edelleen gp340 ja SP-D-surfaktatanttiproteiinin välistä sitoutumista. Vaikka gp340 pystyi sitoutumaan SPD:hen muussakin kuin mannaania sitovassa kohdassa, ei tämä gp340 ja SP-D-sitoutuminen tuottanut kuitenkaan mitään lisäetua antivirusinteraktioihin.
SP-D ja musiini ( mucus) osoittivat interaktiivista yhteistoimintaa ilmeisesti sen takia, että SP-D esti viruksen neuraminidaasia (NA).
Markkinoilla oleva NA-inhibiittori oseltamivir ( Tamiflu) osoitti samankaltaista vaikutusta ja lisäsi myös SP-A surfaktantin (keuhkojen pintajännitystä alentavan tekijän) neutraloivaa toimintaa ja bronkoalveolaarista huuhtelunestettä.
Täten oseltamivir on yhteistyössä luonnollisten immuuniproteiinien kanssa estäessään epiteelisolujen initiaalista infektiota.
LÄHDE:
White MR, Crouch E et.al. Cooperative anti-influenza activities of respiratory innate immune proteins and neuraminidase inhibitor. Am J Physiol Lung Cell Mol Physiol. 2005 May;288(5):L831-40. Epub 2004 Dec 17.
Department of Medicine, Section of Hematology/Oncology, Boston University, School of Medicine, Boston, MA
Terminologia:
surfactant collectins, pintajännitystä alentavat kollektiinit
surfactant proteins A and D (SP-A and D), surfaktanttiproteiinit SP-A ja SP-D
http://ising.phys.cwru.edu/surfactants/proteins.html
scavenger receptor-rich glycoprotein 340 (gp340). glykoproteiini 340, jossa on vapaita radikaaleja pyydystäviä reseptoreita paljon
mucins, musiinit, limaaineet
sialic acid ligands, sialihappoon asettuvia molekyylejä
oseltamivir, neuraminidaseinhibitor, neuraminidaasin estäjä
PIDÄ HUOLTA OMISTA LUONNOLLISISTA SURFAKTANTEISTASI. Tamiflu ja Relenza auttavat ehkä siksi, että lääke tekee yhteistyötä oman puolustuksen keuhkosurfaktanttien ja keuhkomusiinien kanssa ja niiden saaatua apua osetamiviriltä keuhkojen toipuminen influenssasta on parempi.
SURFAKTANTTIEN muodostuminen on parasta, kun on sopivasti ruokaa ja juomaa ja ehkä erityisesti maitotuotteita, kermaa, maitorasvoja, kasvisrasvoja, sipulia, C-vitamiinia, acetylcysteiiniä ( poretablettina käytettävä lääke) . Hyvällä yleiskunnolla ja sopivalla ravinnolla ja antioksidanteilla voi tehostaa luonnollisen puolustuksensa surfaktanttien muodostusta.
On eduksi, jos ei polta tupakkaa.
Influenssan ja kuivan yskän tullen on eduksi käyttää jotain yskänärsytystä hillitsevää lääkettä.
fredag 12 juni 2009
PB2, PB1-F2 ja M2 geenit: Lajirajan ylitys
Pohdinta pandemisesta potentiaalista koskien HN virusgeenejä ja proteiineja PB2, PB1-F2, M2
1. UUTTA TIETOA VUODELTA 2008
MITEN H5N1 PB2-geeni on evoluutiossaan nykyisin?
LÄHDE: Miotto O et al. Identification of human-to-human transmissibility factors in PB2 proteins of influenza A by large-scale mutual information analysis. BMC Bioinformatics. 2008;9 Suppl 1:S18.
Kun taistellaan jotain patogeenia vastaan, on olennaisen tärkeää pystyä identifioimaan niitä mutaatioita, jotka antavat patogeenille ainutlaatuisia ominaisuuksia, kuten kykyä tunkeutua tiettyihin isäntälajeihin.Tässä työssä tutkijat vertailevaa analyysiä käyttämällä kuvaavat erästä uutta metodia, jolla voi identifioida aminohappokohtia, jotka ovat ominaisia spesifisille proteiinisekvensseille. He kehittivät AVANA-software pehmotavaraa. (AVANA software utilizes sequence annotations to select sets for comparison, according to user-specified criteria.) He sovelsivat tätä metodia analysoiden influenza A viruksen PB2- proteiinin sekvenssejä. He identifioivat komponentteja, joissa oli tapahtunut adaptaatio (sopeutuminen) ihmisestä toiseen välittyvään tartuntatapaan ja sitten he rekonstruoivat tapahtuman mutaatiohistorian.
TULOKSET. Tutkijat vertailivat yli 3000 kpl influenssaviruksen PB2 sekvenssiä ihmistartunnoista ja lintuisolaateista. Tässä analyysissä identifioitui 17 karakteristista kohtaa, joista viisi on esiintynyt niissä ihmisiin tarttuneissa kannoissa, joita on ollut olemassa jo vuoden 1918 Espanjan taudista asti. 16 näistä mainituista kohdista sijaitsee funktionaalisissa domaaneissa, mikä viittaa siihen, että niillä voi olla osuutta isäntälajispesifisyydessä. Karakterististen kohtien luetteloa käytettiin seulottaessa esiin sekvenssejä historiallisista isolaateista . Näitä tyyppijälkiä sitten järjestettiin kronologisesti ja havaittiin, miten evoluutionaalisessa aikajanassa PB2 proteiini adaptoituu ihmiseen.
Tämä tutkimus osoittaa, että keskeinen, monipuolinen informaatio on vahva työväline sekvenssisettien vertailevassa analyysissä, kun tällöin on saatavilla täydellisintä selkotietoa niistä funktionaalisista komponenteista, jotka osallistuvat PB2-proteiinin adaptaatioon ihmiskuntaan päin. Tutkijat pystyivät vahvistamaan aiemmin raportoituja löytöjä ja lisäksi monia uusia karakteristisia kohtia H5N1 viruksen PB2:ssa tuli raportoitua. Karakterististen kohtien luetteloa käyttämällä generoituneet sekvenssijäljet luonnehtivat suppeasti yksittäisten virusisolaattien adaptoitumisen tyyppiominaisuuksia. Näistä varhaisista ihmis-influenssavirusten ”jäljistä”selvitetty evolutionaalinen aikajana viittaa siihen, että ominaispiirteiden variantit kehittyvät nopeasti ja pysyvät sitten huomattavan stabiileina kautta myöhempien pandemioitten.
Tämän lisäksi tutkijat sanovat, että ihmisiä infektoivien H5N1 virusisolaattien antamat ”jäljet” viittaavat siihen, että tämä lintuviruksen alatyyppi omaa nykyisellään vain matala-asteisen pandemisen potentiaalin, vaikkakin sillä on enemmän ihmiseen adaptoituneita komponentteja kuin useimmilla muilla lintuvirusalatyypeillä.
In addition, the signatures of human-infecting H5N1 isolates suggest that this avian subtype has low pandemic potential at present, although it presents more human adaptation components than most avian subtypes.
2. TIETOA VUODELTA 2006
Geenien M2 ja PB1-F2 osallistuminen adaptaatioon.
LÄHDE: Smith GJ. Evolution and adaptation of H5N1 influenza virus in avian and human hosts in Indonesia and Vietnam. Virology. 2006 Jul 5;350(2):258-68. Epub 2006 May 19.
Nyt (vuonna 2006) on korkeastipatogeeninen HPAIV H5N1 jo endeemisenä Etelä ja Kaakkois-Aasiassa siipikarjassa ja taudinpurkauksia esiintyy leviävänä Keski-Aasiassa, Euroopassa ja Afrikassa. Lajirajan ylittämisiä tapahtuu jatkuvasti ihmiseen päin- (tosin melkoisen vähäisessä määrin verrattuna populaation totaali määriin) Vietnamissa , Thaimaassa, Kambodjassa, Indonesiassa ja Kiinassa, mikä herättää ajatuksia pandemisesta kehityksestä.
Tutkijaryhmä alkoikin geneettisesti luonnehtia 82 kpl H5N1 virusta, joita oli kerätty siipikarjasta kautta Indonesian ja Vietnamin ja 11 kpl oli eristetty ihmisistä E- Vietnamissa. Yleisiä tietueita hyödynnettiin, jotta saatiin käsitystä näiden virusten tarttumisista, endeemisyydestä ja evoluutiosta. Fylogeneettisesti kaikki indonesialaiset virukset olivat H5N1 Z-genotyyppiä, mikä viittaisi yksittäiseen alkutartuntaan, joka sitten oli levinnyt maassa kauttaaltaan parissa vuodessa.
Jatkuvat virusaktiviteetit Indonesiassa johtuvat lähinnä siipikarjalintujen siirtelemisestä maan rajojen sisällä ennemminkin kuin muuttolintujen liikehdinnästä. Indonesiassa ja Vietnamissa H5N1 virukset ovat tehneet evoluutionsa geograafisesti erotettaviin ryhmiin kummassakin maassa .
Kun analysoitiin H5N1- viruksen genotyypin Z-genomia, osoittautui, että vain M2 ja PB1-F2 geenit olivat positiivisesti valiutuvia, mikä viittaa siihen, että nämä geenit osallistuisivat tämän viruksen adaptaatioihin uusiin isäntiin lajirajan ylittävissä transmissioissa. Aminohappotasossa 12 aminohappoa oli positiivisesti valiutuvia näissä genotyyppi Z- viruksissa HA ja PB1-F2 proteiineissa. Muutamia näistä aminohapoista oli useammin tavattavissa ihmisistä eristetyissä viruksissa kuin linnuista eristetyissä ja ne korreloivat viruksen antigeenisyyteen ja reseptoriin sitoutumiseen. Tutkimuksessa saatiin oivallusta H5N1-viruksen käynnissä olevasta evoluutiosta sen välittyessä eri lintulajeihin ja myös lajirajan ylittämisiin linnuista ihmis-isäntiin päin.
3. TIETOA VUODELTA 2004
H5N1 genotyyppi Z
H5N1 genotyppi Z oli se korkeasti patogeeni (HPAIV) H5N1, joka riehui Kiinassa ja seitsemässä muussa maassa vuosina 2003-4. Nämä maat olivat Japani, E-Korea, Vietnam, Laos, Kambodja ja Indonesia. Tämä virus tappoi siipikarjaa miljoonittain, mutta myös välittyi ihmisiin ollen 54 % letaali infektoituneissa. Näistä ajoista genotyyppi Z alkoi olla dominantti Aasiassa. Tämä havaittiin ensiksi marraskuussa 2002 Hongkongissa ja se oli antigeenisesti erilainen kuin se virus, jonka havaittiin olleen letaali vesilinnuille 1997-2002.
Ihmisiä ja siipikarjaa vuonna 2004 Aasiassa infektoinut virus oli ”Z- genotyypin variantti”. Muuttolinnut olivat ilmeisesti siirtäneet virusta lopulta kautta Aasian.
LÄHDE:
Webster RG, The spread of the H5N1 bird flu epidemic in Asia in 2004. Arch Virol Suppl. 2005;(19):117-29.
Li KS. Genesis of a highly pathogenic and potentially pandemic H5N1 influenza virus in eastern Asia. Nature. 2004 Jul 8;430(6996):209-13.
4. MIGRATIONMAP
http://www.newsdesk.umd.edu/images/flumigrationmap.gif
5. KUVA VIRUKSEN 8 GEENISTÄ
http://www.nature.com/nrmicro/journal/v2/n10/images/nrmicro979-f
6. KUVA viruksen GEENITUOTTEISTA, PROTEIINEISTA
http://www.aasv.org/shap/issues/v13n3/v13n3p157t1.gif
http://content.answers.com/main/content/wp/en-commons/0/02/Flu_und_legende_color_c.jpg
20/03/2008 16:17 Pandemisuuspotentiaalin pohdintaa L Bright
1. UUTTA TIETOA VUODELTA 2008
MITEN H5N1 PB2-geeni on evoluutiossaan nykyisin?
LÄHDE: Miotto O et al. Identification of human-to-human transmissibility factors in PB2 proteins of influenza A by large-scale mutual information analysis. BMC Bioinformatics. 2008;9 Suppl 1:S18.
Kun taistellaan jotain patogeenia vastaan, on olennaisen tärkeää pystyä identifioimaan niitä mutaatioita, jotka antavat patogeenille ainutlaatuisia ominaisuuksia, kuten kykyä tunkeutua tiettyihin isäntälajeihin.Tässä työssä tutkijat vertailevaa analyysiä käyttämällä kuvaavat erästä uutta metodia, jolla voi identifioida aminohappokohtia, jotka ovat ominaisia spesifisille proteiinisekvensseille. He kehittivät AVANA-software pehmotavaraa. (AVANA software utilizes sequence annotations to select sets for comparison, according to user-specified criteria.) He sovelsivat tätä metodia analysoiden influenza A viruksen PB2- proteiinin sekvenssejä. He identifioivat komponentteja, joissa oli tapahtunut adaptaatio (sopeutuminen) ihmisestä toiseen välittyvään tartuntatapaan ja sitten he rekonstruoivat tapahtuman mutaatiohistorian.
TULOKSET. Tutkijat vertailivat yli 3000 kpl influenssaviruksen PB2 sekvenssiä ihmistartunnoista ja lintuisolaateista. Tässä analyysissä identifioitui 17 karakteristista kohtaa, joista viisi on esiintynyt niissä ihmisiin tarttuneissa kannoissa, joita on ollut olemassa jo vuoden 1918 Espanjan taudista asti. 16 näistä mainituista kohdista sijaitsee funktionaalisissa domaaneissa, mikä viittaa siihen, että niillä voi olla osuutta isäntälajispesifisyydessä. Karakterististen kohtien luetteloa käytettiin seulottaessa esiin sekvenssejä historiallisista isolaateista . Näitä tyyppijälkiä sitten järjestettiin kronologisesti ja havaittiin, miten evoluutionaalisessa aikajanassa PB2 proteiini adaptoituu ihmiseen.
Tämä tutkimus osoittaa, että keskeinen, monipuolinen informaatio on vahva työväline sekvenssisettien vertailevassa analyysissä, kun tällöin on saatavilla täydellisintä selkotietoa niistä funktionaalisista komponenteista, jotka osallistuvat PB2-proteiinin adaptaatioon ihmiskuntaan päin. Tutkijat pystyivät vahvistamaan aiemmin raportoituja löytöjä ja lisäksi monia uusia karakteristisia kohtia H5N1 viruksen PB2:ssa tuli raportoitua. Karakterististen kohtien luetteloa käyttämällä generoituneet sekvenssijäljet luonnehtivat suppeasti yksittäisten virusisolaattien adaptoitumisen tyyppiominaisuuksia. Näistä varhaisista ihmis-influenssavirusten ”jäljistä”selvitetty evolutionaalinen aikajana viittaa siihen, että ominaispiirteiden variantit kehittyvät nopeasti ja pysyvät sitten huomattavan stabiileina kautta myöhempien pandemioitten.
Tämän lisäksi tutkijat sanovat, että ihmisiä infektoivien H5N1 virusisolaattien antamat ”jäljet” viittaavat siihen, että tämä lintuviruksen alatyyppi omaa nykyisellään vain matala-asteisen pandemisen potentiaalin, vaikkakin sillä on enemmän ihmiseen adaptoituneita komponentteja kuin useimmilla muilla lintuvirusalatyypeillä.
In addition, the signatures of human-infecting H5N1 isolates suggest that this avian subtype has low pandemic potential at present, although it presents more human adaptation components than most avian subtypes.
2. TIETOA VUODELTA 2006
Geenien M2 ja PB1-F2 osallistuminen adaptaatioon.
LÄHDE: Smith GJ. Evolution and adaptation of H5N1 influenza virus in avian and human hosts in Indonesia and Vietnam. Virology. 2006 Jul 5;350(2):258-68. Epub 2006 May 19.
Nyt (vuonna 2006) on korkeastipatogeeninen HPAIV H5N1 jo endeemisenä Etelä ja Kaakkois-Aasiassa siipikarjassa ja taudinpurkauksia esiintyy leviävänä Keski-Aasiassa, Euroopassa ja Afrikassa. Lajirajan ylittämisiä tapahtuu jatkuvasti ihmiseen päin- (tosin melkoisen vähäisessä määrin verrattuna populaation totaali määriin) Vietnamissa , Thaimaassa, Kambodjassa, Indonesiassa ja Kiinassa, mikä herättää ajatuksia pandemisesta kehityksestä.
Tutkijaryhmä alkoikin geneettisesti luonnehtia 82 kpl H5N1 virusta, joita oli kerätty siipikarjasta kautta Indonesian ja Vietnamin ja 11 kpl oli eristetty ihmisistä E- Vietnamissa. Yleisiä tietueita hyödynnettiin, jotta saatiin käsitystä näiden virusten tarttumisista, endeemisyydestä ja evoluutiosta. Fylogeneettisesti kaikki indonesialaiset virukset olivat H5N1 Z-genotyyppiä, mikä viittaisi yksittäiseen alkutartuntaan, joka sitten oli levinnyt maassa kauttaaltaan parissa vuodessa.
Jatkuvat virusaktiviteetit Indonesiassa johtuvat lähinnä siipikarjalintujen siirtelemisestä maan rajojen sisällä ennemminkin kuin muuttolintujen liikehdinnästä. Indonesiassa ja Vietnamissa H5N1 virukset ovat tehneet evoluutionsa geograafisesti erotettaviin ryhmiin kummassakin maassa .
Kun analysoitiin H5N1- viruksen genotyypin Z-genomia, osoittautui, että vain M2 ja PB1-F2 geenit olivat positiivisesti valiutuvia, mikä viittaa siihen, että nämä geenit osallistuisivat tämän viruksen adaptaatioihin uusiin isäntiin lajirajan ylittävissä transmissioissa. Aminohappotasossa 12 aminohappoa oli positiivisesti valiutuvia näissä genotyyppi Z- viruksissa HA ja PB1-F2 proteiineissa. Muutamia näistä aminohapoista oli useammin tavattavissa ihmisistä eristetyissä viruksissa kuin linnuista eristetyissä ja ne korreloivat viruksen antigeenisyyteen ja reseptoriin sitoutumiseen. Tutkimuksessa saatiin oivallusta H5N1-viruksen käynnissä olevasta evoluutiosta sen välittyessä eri lintulajeihin ja myös lajirajan ylittämisiin linnuista ihmis-isäntiin päin.
3. TIETOA VUODELTA 2004
H5N1 genotyyppi Z
H5N1 genotyppi Z oli se korkeasti patogeeni (HPAIV) H5N1, joka riehui Kiinassa ja seitsemässä muussa maassa vuosina 2003-4. Nämä maat olivat Japani, E-Korea, Vietnam, Laos, Kambodja ja Indonesia. Tämä virus tappoi siipikarjaa miljoonittain, mutta myös välittyi ihmisiin ollen 54 % letaali infektoituneissa. Näistä ajoista genotyyppi Z alkoi olla dominantti Aasiassa. Tämä havaittiin ensiksi marraskuussa 2002 Hongkongissa ja se oli antigeenisesti erilainen kuin se virus, jonka havaittiin olleen letaali vesilinnuille 1997-2002.
Ihmisiä ja siipikarjaa vuonna 2004 Aasiassa infektoinut virus oli ”Z- genotyypin variantti”. Muuttolinnut olivat ilmeisesti siirtäneet virusta lopulta kautta Aasian.
LÄHDE:
Webster RG, The spread of the H5N1 bird flu epidemic in Asia in 2004. Arch Virol Suppl. 2005;(19):117-29.
Li KS. Genesis of a highly pathogenic and potentially pandemic H5N1 influenza virus in eastern Asia. Nature. 2004 Jul 8;430(6996):209-13.
4. MIGRATIONMAP
http://www.newsdesk.umd.edu/images/flumigrationmap.gif
{kind=link}
5. KUVA VIRUKSEN 8 GEENISTÄ
http://www.nature.com/nrmicro/journal/v2/n10/images/nrmicro979-f
6. KUVA viruksen GEENITUOTTEISTA, PROTEIINEISTA
http://www.aasv.org/shap/issues/v13n3/v13n3p157t1.gif
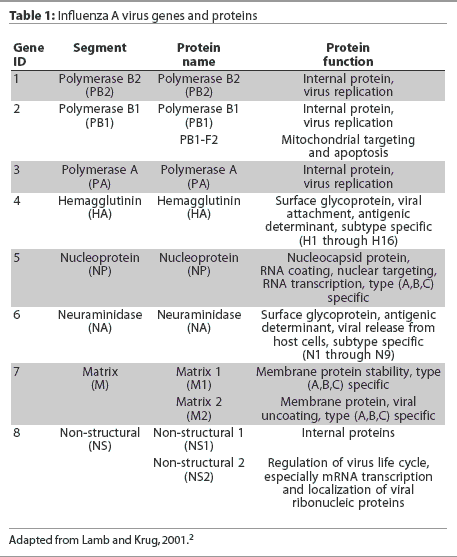{kind=link}
http://content.answers.com/main/content/wp/en-commons/0/02/Flu_und_legende_color_c.jpg
{kind=link}
20/03/2008 16:17 Pandemisuuspotentiaalin pohdintaa L Bright
A (H1N1) influenssaviruksen genomi
Olen vuosien aikana katsonut tätä influenssaviruksen genomia useankin kerran. Jollain tavalla minusta välillä tuntuu epäoikeudenmukaiseltakin, että juuri tästä genomista on valtava tietämys ja siitä on suunnaton tutkimus menossa, ja samaan aikaan jokin klassisesti tuhansia , tai satoja tuhansia tai miljoonia ihmisiä totunnaisesti aina tappava virus jatkaa samoja ratojaan aivan kuin tämän influenssaviruksen suuressa varjossa aivan huomaamatta, eikä niille lukuisille fataaleille viruksille liikene sitten kansainvälisiä resursseja esim. rokotekehittelyyn tarpeeksi, kun influenssa-asia nielee kuin haikala kaikki varat.
Toisaalta asia on käsitettävä, koska harva virus kykenee olemaan potentiaalisti niin paha kuin influenssavirus kaikkein pahimmassa muodosaan voisi olla. Preventiivisesti ottaen asia on kyllä oikeudenmukaista. ESIM. Ebola on kyllä paha ja tappava, mutta se on endeeminen. Influenssavirus taas voi päästä aivan kaikkiin ilmasto-olosuhteisiin maapallolla kunnioittamatta mitään endeemisyyden ideaakaan- ellei nyt sitten reservoaarina.
Sitten toinen asia, vaikuttaa siltä, että influenssaa saa mikä tahansa elävä. Kohta voi jo odottaa että kukkakin aivastaa. Tarkoitan, että ehkä kasveissakin voi olla tätä virusta , ainakin kuin reservoaarissa, jos se oikein yrittää kehittyä. Mutta ihminen, ihmiskunta, on sen genomille täysi välttämättömyys. Sen genomin täytyy mennä ihmisen DNA genomin viereen ottamaan mallia ja sitten replikoitumaan. Vasta kun se on kehrännyt maapallolta parin kolmen pandemisen aallon kanssa tarpeeksi materiaalia genominsa piristykseksi, se katoaa jonnekin taas. Sen aivotrusti katoaa jonnekin. Siinä on jotain hämähäkkimäisen tapaista katoamista maisemasta yhtäkkiä.
Tämä virus intelligenssi on kiinnostava,voi sanoa. Viruksissa on jotain hienoa. Ainakin se, että ne panevat ihmiset ajattelemaan enemmän. Viruksista ihminen kehittyy ja kehittää tietokoneensa ja globaalin kommunikaationsa aivan huippuunsa.
INFLUENSSA A viruksen genomissa on 8 segmenttiä ja ne esiintyvät erillisinä. Virus pakkaa joka palan erikseen hyvin täydellisesti. Jokainen geeni on RNA-rihma, negatiivinen yksittäinen rihma, joka on nukleotidiketju, jonka 5´ ja 3´ pää on kiinnitetty polymeraasikompleksiin, aivan kuin helminauha kiinnitetään lukkoonsa. Rihmassa on NP (nukleoproteiini) "helmiä" ja rihma kiertää näitä NP-helmien nauhaa helixmäisesti.
( Kahdeksan segmenttiä: PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS)
Tämä helminauha ei esiinny aukinaisena ympyränä ikäänkuin olisi kaulassa, vaan ikäänkuin pienimpään tilavuuteen helixmäisesti kiertyneenä, mitä nyt noilta NP-helmiltä saa kiertymää aikaan. Tämä on vRNP partikkeli nimeltään. itse asiassa se näyttää hieman sauvamaiselta.
Toiseen päätyyn kuitenkin muodosotuu pieni rinkula.
"Lukon"lähellä olevassa kompleksissa on entsyymejä ä PB2, PB1 ja PA valmiina proteiinina ns. polymeraasikompleksina, eväänä mukana äiti-virukselta ja vRNA 5´-pää kiinnittyy PB2 ja NP proteiiniin, kuin tosiaan helminauhan lukkoon.
http://www.nature.com/emboj/journal/v17/n19/images/7591278f8.jpg
Helminauha voi tietysti aueta, kun on aika toimia ja nauha liukuu kuin lukkoalueen entsyymirenkaan läpi kuin kohesiinirenkaassa kai.
Polymeraasipaketti järjestää siten, että entsyymit napsaavat solun mRNA:lta alukkeen (CAP-primer), leikkavat sen irti ja kiinnittävät sen helminauhaketjussa olevan RNA:n 3´-päähän ja siitä sitten onkin hyvä lähteä tikittämään mRNA molekyyliä polymeraaseilla esiin cis-asemasta ja peräleijaksi asetetaan polyadenylaatiota lähettiRNA:n elämän pitkittämiseksi. Samalla pääsee RNA ikäänkuin oikenemaan mikä varmasti provosoi kaikenlaista mishmatch etc korjaantumista jokaiseen nukleotidiin ja "vironolosuhteiden nälkäajan ketju" rikastuu ja priistyy tumanesteissä ja sitten voi tuottua myös antigenomi, tässä tapauksessa (+) cRNA:n.
Kun virus on saanut pukattua omaa mRNA:ta tekemään uusia alkuproteiineja, niitä alkaa virrata tuma-alueelle ja cRNA alkaa trans-asemasta tuottaa sitten uutta virionia ihka tuoreilla polymeraasi-entsyymeillä. ( Tietysti toiset mRNA:t tuottavat myös runsaasti sytoplasmaan loppuvaiheen tuotteita, jotka lähetetään solukalvoon tekemään pesää uusille viruspartikkeleille, sitä virionkapselin massiivaa; M proteiinia, HA, NA proteiineja)
Apurina replikaation ja transkription alalla toimii NS, jolla onkin paljon tekemistä. Sen tulee katsoa, että missään vaiheessa virusRNA, vaikka juosteet olisivatvRNP partikkelissa lähekkäin, lähellä, ei anna dsRNA signaalia. NS leipoutuu vRNP partikkeleihin tuollaisissa strategisissa kohdissa ja niin solun virussensorit eivät huomaa ollenkaan viruksen olemassa oloa ja se pääsee kuin näkymättömänä tumaan aivan lojaalia tietä, tumakalvon aukoista, poruksista, itse solun importiinien avulla, sillä NS kiinnittää myös importiineja käsittäen NLS ja NoLS kohtia, jotka vievät sitä suoraan tumaan ja tumajyväseen.
Kun virus syntetisoi monipuolisesti genomia ja antigenomia tulee välillä dsRNA muotoa, jonka sitten NS-kiinnittää itseeensä "neutraloi", eikä Interferonijärjestelmä pysty heräämään. Interferonit heräisivät dsRN:sta ja herättäisivät sata antiviraalia geeniä. Siis itse asiassa solun mahtavan tehokas DNA genomi nukahtaa ikuiseen uneen ja säätyy nopeasti aivan kokonaan hajoamaan samalla, kun sitten solun integraatio kaikin puolin hajoaa ja uudet virionit lähtevät eteenpäin valmistettua tietä.
Jokainen virusgeenisegmentti vaikuttaa apoptoottisesti ihmisDNA:han, toiset enemmän toiset vähemmän. Yksi geeneistä on oikein erikoistunut ihmissolun mitokondrian hävittämiseen. (PB1-F2) .'
Eri virus make-upit , mitä influenssa virus esittää ovat eri asteisesti virulentteja tämän seikan suhteen. Virulenssin määrääviä seikkoja on monta, esim tapa jolla virus kokoaa 8 segmenttiään. Virulenssiin vaikuttaa jos se on yhtä ja samaa alkuperää esim kokonaan lintuvirus. Ne ovat kovin apoptoosia aiheuttavia geenisegmenttejä kaikki.
Tällä hetkellä on lintuviruksesta tullut vain kaksi geenisegmenttiä tähän uuteen pandemiseen virukseen: PB2 ja PA: Helminauhan lukko-osan tekijöitä molemmat.
Samoin ihmisperäinen PB1 kuuluu siihen helminauhan lukkoon.
Kaikki massiiviset rakenteelliset ja nonstrukturellit proteiinit ovat eri sikaviruksista, joten ihmiskeho on sen massiivalle vastaanottavainen. (HA, NP, NA, M, NS)
PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS
Toisaalta asia on käsitettävä, koska harva virus kykenee olemaan potentiaalisti niin paha kuin influenssavirus kaikkein pahimmassa muodosaan voisi olla. Preventiivisesti ottaen asia on kyllä oikeudenmukaista. ESIM. Ebola on kyllä paha ja tappava, mutta se on endeeminen. Influenssavirus taas voi päästä aivan kaikkiin ilmasto-olosuhteisiin maapallolla kunnioittamatta mitään endeemisyyden ideaakaan- ellei nyt sitten reservoaarina.
Sitten toinen asia, vaikuttaa siltä, että influenssaa saa mikä tahansa elävä. Kohta voi jo odottaa että kukkakin aivastaa. Tarkoitan, että ehkä kasveissakin voi olla tätä virusta , ainakin kuin reservoaarissa, jos se oikein yrittää kehittyä. Mutta ihminen, ihmiskunta, on sen genomille täysi välttämättömyys. Sen genomin täytyy mennä ihmisen DNA genomin viereen ottamaan mallia ja sitten replikoitumaan. Vasta kun se on kehrännyt maapallolta parin kolmen pandemisen aallon kanssa tarpeeksi materiaalia genominsa piristykseksi, se katoaa jonnekin taas. Sen aivotrusti katoaa jonnekin. Siinä on jotain hämähäkkimäisen tapaista katoamista maisemasta yhtäkkiä.
Tämä virus intelligenssi on kiinnostava,voi sanoa. Viruksissa on jotain hienoa. Ainakin se, että ne panevat ihmiset ajattelemaan enemmän. Viruksista ihminen kehittyy ja kehittää tietokoneensa ja globaalin kommunikaationsa aivan huippuunsa.
INFLUENSSA A viruksen genomissa on 8 segmenttiä ja ne esiintyvät erillisinä. Virus pakkaa joka palan erikseen hyvin täydellisesti. Jokainen geeni on RNA-rihma, negatiivinen yksittäinen rihma, joka on nukleotidiketju, jonka 5´ ja 3´ pää on kiinnitetty polymeraasikompleksiin, aivan kuin helminauha kiinnitetään lukkoonsa. Rihmassa on NP (nukleoproteiini) "helmiä" ja rihma kiertää näitä NP-helmien nauhaa helixmäisesti.
( Kahdeksan segmenttiä: PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS)
Tämä helminauha ei esiinny aukinaisena ympyränä ikäänkuin olisi kaulassa, vaan ikäänkuin pienimpään tilavuuteen helixmäisesti kiertyneenä, mitä nyt noilta NP-helmiltä saa kiertymää aikaan. Tämä on vRNP partikkeli nimeltään. itse asiassa se näyttää hieman sauvamaiselta.
Toiseen päätyyn kuitenkin muodosotuu pieni rinkula.
"Lukon"lähellä olevassa kompleksissa on entsyymejä ä PB2, PB1 ja PA valmiina proteiinina ns. polymeraasikompleksina, eväänä mukana äiti-virukselta ja vRNA 5´-pää kiinnittyy PB2 ja NP proteiiniin, kuin tosiaan helminauhan lukkoon.
http://www.nature.com/emboj/journal/v17/n19/images/7591278f8.jpg
{kind=link}
Helminauha voi tietysti aueta, kun on aika toimia ja nauha liukuu kuin lukkoalueen entsyymirenkaan läpi kuin kohesiinirenkaassa kai.
Polymeraasipaketti järjestää siten, että entsyymit napsaavat solun mRNA:lta alukkeen (CAP-primer), leikkavat sen irti ja kiinnittävät sen helminauhaketjussa olevan RNA:n 3´-päähän ja siitä sitten onkin hyvä lähteä tikittämään mRNA molekyyliä polymeraaseilla esiin cis-asemasta ja peräleijaksi asetetaan polyadenylaatiota lähettiRNA:n elämän pitkittämiseksi. Samalla pääsee RNA ikäänkuin oikenemaan mikä varmasti provosoi kaikenlaista mishmatch etc korjaantumista jokaiseen nukleotidiin ja "vironolosuhteiden nälkäajan ketju" rikastuu ja priistyy tumanesteissä ja sitten voi tuottua myös antigenomi, tässä tapauksessa (+) cRNA:n.
Kun virus on saanut pukattua omaa mRNA:ta tekemään uusia alkuproteiineja, niitä alkaa virrata tuma-alueelle ja cRNA alkaa trans-asemasta tuottaa sitten uutta virionia ihka tuoreilla polymeraasi-entsyymeillä. ( Tietysti toiset mRNA:t tuottavat myös runsaasti sytoplasmaan loppuvaiheen tuotteita, jotka lähetetään solukalvoon tekemään pesää uusille viruspartikkeleille, sitä virionkapselin massiivaa; M proteiinia, HA, NA proteiineja)
Apurina replikaation ja transkription alalla toimii NS, jolla onkin paljon tekemistä. Sen tulee katsoa, että missään vaiheessa virusRNA, vaikka juosteet olisivatvRNP partikkelissa lähekkäin, lähellä, ei anna dsRNA signaalia. NS leipoutuu vRNP partikkeleihin tuollaisissa strategisissa kohdissa ja niin solun virussensorit eivät huomaa ollenkaan viruksen olemassa oloa ja se pääsee kuin näkymättömänä tumaan aivan lojaalia tietä, tumakalvon aukoista, poruksista, itse solun importiinien avulla, sillä NS kiinnittää myös importiineja käsittäen NLS ja NoLS kohtia, jotka vievät sitä suoraan tumaan ja tumajyväseen.
Kun virus syntetisoi monipuolisesti genomia ja antigenomia tulee välillä dsRNA muotoa, jonka sitten NS-kiinnittää itseeensä "neutraloi", eikä Interferonijärjestelmä pysty heräämään. Interferonit heräisivät dsRN:sta ja herättäisivät sata antiviraalia geeniä. Siis itse asiassa solun mahtavan tehokas DNA genomi nukahtaa ikuiseen uneen ja säätyy nopeasti aivan kokonaan hajoamaan samalla, kun sitten solun integraatio kaikin puolin hajoaa ja uudet virionit lähtevät eteenpäin valmistettua tietä.
Jokainen virusgeenisegmentti vaikuttaa apoptoottisesti ihmisDNA:han, toiset enemmän toiset vähemmän. Yksi geeneistä on oikein erikoistunut ihmissolun mitokondrian hävittämiseen. (PB1-F2) .'
Eri virus make-upit , mitä influenssa virus esittää ovat eri asteisesti virulentteja tämän seikan suhteen. Virulenssin määrääviä seikkoja on monta, esim tapa jolla virus kokoaa 8 segmenttiään. Virulenssiin vaikuttaa jos se on yhtä ja samaa alkuperää esim kokonaan lintuvirus. Ne ovat kovin apoptoosia aiheuttavia geenisegmenttejä kaikki.
Tällä hetkellä on lintuviruksesta tullut vain kaksi geenisegmenttiä tähän uuteen pandemiseen virukseen: PB2 ja PA: Helminauhan lukko-osan tekijöitä molemmat.
Samoin ihmisperäinen PB1 kuuluu siihen helminauhan lukkoon.
Kaikki massiiviset rakenteelliset ja nonstrukturellit proteiinit ovat eri sikaviruksista, joten ihmiskeho on sen massiivalle vastaanottavainen. (HA, NP, NA, M, NS)
PB2/ PB1/ PA/ HA/ NP/ NA/ M/ NS
Sikainfluenssavirukset syyniin
Siihen aikaan kun WHO nosti 11.5.2009 pandemisen hälytyksen tasoon 5 ...
LÄHDE: Smith GJ, Vijaykrishna D,et al. Origins and evolutionary genomics of the 2009 swine-origin H1N1 influenza A epidemic. State Key Laboratory of Emerging Infectious Diseases & Department of Microbiology, Hong Kong SAR, China. Nature. 2009 Jun 11
Tällä S-OIV ( swine-origin influenza A) viruksella A(H1N1) on kykyä kehittyä tämän vuosituhannen ensimmäiseksi pandemiaksi ( vakuuttivat keväällä tutkijat jo ennen kuin WHO julisti pandemian alkaneeksi)
Maaliskuussa ja varhain huhtikuussa 2009 oli puhjennut ihmisissä uusi sikavirusalkuperäinen(S-OIV) A influenssavirus (H1N1) Meksikossa ja USA:ssa. Ensimmäisten viikkojen havaintojen mukaan virus levisi maailmanlaajuisesti 30 maahan toukokuun 11. päivään mennessä siirtymällä ihmisestä ihmiseen, jolloin WHO nosti pandemisen hälytystason (6-numeroisessa asteikossa) tasoon 5.
Tutkijat sanovat katsauksessaan:
Olemme tehneet EVOLUTIONAALISEN analyysin arvioiden S-OIV-epidemian alkuperän aikajanaa ja varhaiskehitystä. Me osoitamme, että se on johtunut useista sioissa kiertäneistä viruksista ja että varhaisin siirtyminen ihmisiin on tapahtunut useita kuukausia ennen kuin purkaus havaittiin
FYLOGENEETTINEN arvio viruksen geneettisistä elossapysymisaukoista viittaa siihen, että on esiintynyt pitkäjaksoinen tutkimattomien esivirusten linja ennen S-OIV purkausta, mikä viittaa sikalinjan virusten uudelleen järjestymiseen (REASSORTEMENT) . Se lie tapahtunut vuosia ennen kuin ihmisissä alkoi infektion leviäminen. Monipuolinen geneettinen esikantajoukko, joka S-OIV:llä on, ei ole keinotekoisesta lähteestä ( esim. rokotteista karannutta)
Koska epidemian historian ajalta ei ole mitään n virunäytteitä , niin geneettisesti kaikkein lähinnä toisiaan olevien sikavirusten luonne ja paikallistuminen selventää tuskin mitään epidemian välittömästä alkuperästä (ORIGIN) , siitä huolimatta, vaikka paneliin sijoitettiin hyvin lähisukulaisia ja ja aiemmin julkaisemattomia sika-influenssa isolaatteja.
Tutkijoitten tuloksista nähdään tarve kartoittaa sikainfluenssaa systemaattisesti ja samoin tarve saada näyttöä uuden geneettisen aineksen siassa sekoittumisesta, mikä voi johtaa ihmisille pandemisten virusten esiintuloon- myös tulevaisuudessa.
12.6.2009 15:08
LÄHDE: Smith GJ, Vijaykrishna D,et al. Origins and evolutionary genomics of the 2009 swine-origin H1N1 influenza A epidemic. State Key Laboratory of Emerging Infectious Diseases & Department of Microbiology, Hong Kong SAR, China. Nature. 2009 Jun 11
Tällä S-OIV ( swine-origin influenza A) viruksella A(H1N1) on kykyä kehittyä tämän vuosituhannen ensimmäiseksi pandemiaksi ( vakuuttivat keväällä tutkijat jo ennen kuin WHO julisti pandemian alkaneeksi)
Maaliskuussa ja varhain huhtikuussa 2009 oli puhjennut ihmisissä uusi sikavirusalkuperäinen(S-OIV) A influenssavirus (H1N1) Meksikossa ja USA:ssa. Ensimmäisten viikkojen havaintojen mukaan virus levisi maailmanlaajuisesti 30 maahan toukokuun 11. päivään mennessä siirtymällä ihmisestä ihmiseen, jolloin WHO nosti pandemisen hälytystason (6-numeroisessa asteikossa) tasoon 5.
Tutkijat sanovat katsauksessaan:
Olemme tehneet EVOLUTIONAALISEN analyysin arvioiden S-OIV-epidemian alkuperän aikajanaa ja varhaiskehitystä. Me osoitamme, että se on johtunut useista sioissa kiertäneistä viruksista ja että varhaisin siirtyminen ihmisiin on tapahtunut useita kuukausia ennen kuin purkaus havaittiin
FYLOGENEETTINEN arvio viruksen geneettisistä elossapysymisaukoista viittaa siihen, että on esiintynyt pitkäjaksoinen tutkimattomien esivirusten linja ennen S-OIV purkausta, mikä viittaa sikalinjan virusten uudelleen järjestymiseen (REASSORTEMENT) . Se lie tapahtunut vuosia ennen kuin ihmisissä alkoi infektion leviäminen. Monipuolinen geneettinen esikantajoukko, joka S-OIV:llä on, ei ole keinotekoisesta lähteestä ( esim. rokotteista karannutta)
Koska epidemian historian ajalta ei ole mitään n virunäytteitä , niin geneettisesti kaikkein lähinnä toisiaan olevien sikavirusten luonne ja paikallistuminen selventää tuskin mitään epidemian välittömästä alkuperästä (ORIGIN) , siitä huolimatta, vaikka paneliin sijoitettiin hyvin lähisukulaisia ja ja aiemmin julkaisemattomia sika-influenssa isolaatteja.
Tutkijoitten tuloksista nähdään tarve kartoittaa sikainfluenssaa systemaattisesti ja samoin tarve saada näyttöä uuden geneettisen aineksen siassa sekoittumisesta, mikä voi johtaa ihmisille pandemisten virusten esiintuloon- myös tulevaisuudessa.
12.6.2009 15:08
WHO julisti H1N1 pandemiseksi (6) 11.6.2009
http://www.who.int/en/
12.6.2006
WHO:n sanoma 11.kesäkuuta
Suomennosta: LÄHDE: Statement to the press by WHO Director-General Dr Margaret Chan
11 June 2009 World now at the start of 2009 influenza pandemic
"Hyvät Naiset ja Herrat,
Huhtikuun lopussa WHO julisti hälytystilan uuden A influenssaviruksen takia.Tämä erityinen H1N1 kanta ei ole aiemmin kiertänyt ihmiskuntaa. Virus on aivan uusi. Virus on tarttuvainen ja leviää helposti ihmisestä toiseen ja maasta toiseen. Tänään on noin 30 000 varmistettua tapausta raportoitu yhteensä 74 eri maasta. Tässä on vain osa asiaa. Muutamaa poikkeusta huomioonottamatta niissä maissa, joissa on esiintynyt suuri määrä sairaustapauksia tilanteen valvonta ja näytteitten otto on hyvä. Leviämistä maasta toiseen ei enää voida jäljittää selkeästi ihmisten tartuntaketjua määrittämällä. Jatkoleviäminen on katsottava väistämättömäksi tapahtumaksi. Olen neuvotellut johtavien influenssa-asiantuntijoiden, virologien ja kansanterveysviranomaisten kanssa. Kansainvälisten terveyttä koskevien säädösten (International Health Regulations) käynnistämien proseduurien mukaisesti olen konsultoinut Poikkeustilakomiteaa (Emergency Committee) , joka on tätä tarkoitusta varten perustettu. Saatavilla olevan näytön ja niihin tutustuneitten asiantunijoitten en arvion perusteella on influenssapandemian tieteelliset kriteerit täyttyneet
Olen sen takia päättänyt kohottaa influenssapandemian hälytystasolta (5) pandemiatasoon (6). Nyt 11. kesäkuuta 2009 maailma on vuoden 2009 influenssapandemiansa alussa. (START).
Olemme nyt pandemian varhaispäivissä Virus leviää tiukan ja huolellisen kontrollin alaisena. EI KOSKAAN ole havaittu pandemiaa näin varhain eikä näin tarkasti seurattu juuri tapahtuvalla ajalla aivan varhaisista alkuhetkistä alkaen. Nyt maailmassa saadaan niittää pandemiavalmiuden etuja, joihin ollaan investoitu viime kuluneen viiden vuoden aikana. Meillä on etumatkaa. Tämä antaa meille vahvan aseman. Mutta samalla se haastaa meitä tietojemme ollessa rajallisia ja tieteellisen epävarmuuden vallitessa saamaan lisää neuvoja ja uusia vakuuksia
Hyvän monitoroinnin , perusteellisten tutkimusten ja eri maitten suoran raportoinnin ansiosta olemme saaneet muutamia varhaisia pikakäsityksiä viruksen leviämisestä ja sen mahdollisesti aiheuttamasta tautikirjosta.
Tiedämme myös , että varhaisen siellä täällä esiintyvän viruksen tavat voivat muuttua äkisti. Virus tekee säännöt ja tämäkin H1N1 kaikkien influenssavirusten tapaan voi muuttaa sääntöjään sen suuremmitta sävelittä milloin tahansa.
Globaalisti ottaen meillä on hyvät syymme uskoa tämän pandemian ainakin varhaispäivinään pysyttelevän asteeltaan kohtalaisen vakavana. Kokemuksesta tiedämme vakavuusasteen vaihtelevan maasta toiseen monista tekijöistä johtuen. Nykytietämyksen mukaan potilaitten valtaenemmistöllä on ollut lieviä oireita ja toipuminen on ollut nopeaa ja täydellistä usein ilman mitään lääkehoitoakin.
Kautta mailman on menehtyneitten luku ollut pieni. Joka ainoa näistä kuolemista on kuitenkin traaginen ja meidän on koottava voimia kohdatessamme niitä yhä lisää. Kuitenkaan emme odota mitään vaikeiden tai fataalien infektioiden äkillistä ja kohtalokasta lisääntymistä.
Tiedämme uuden H1N1 viruksen infektoivan nuorta väkeä. Miltei kaikilla laajemman ja pitkäaikaisemman purkauksen alueilla pääosa infektoituneesta väestöstä on ollut alle 25 vuotiaita.
Joissain maissa noin 2% infektoituneista on saanut vakavamman taudin, jossa esiintyy nopea paheneminen kohti henkeä uhkaavaa keuhkokuumetta.
Useimmat vaikeat tai fataalit infektiot ovat kohdanneet aikuisia, jotka ovat 30-50 vuoden ikäisiä.
Tällainen tautimalli on merkitsevästi kausiluontoisesta influenssaepidemiasta poikkeava, missä useimmat menehtymiset kohtaavat iäkästä väestöä.
Monesti, tosin ei jokaisessa tapauksessa, vakava sairastuminen on tapahtunut siinä väestössä, jolla on jokin krooninen perussairaus . Alustaviin rajallisiin tietoihin pohjautuen voidaan sanoa, että näihin sairauksiin kuuluu hengitysteiden taudit, varsinkin astma, sydän-ja verenkiertoelinten taudit, diabetes, autoimmuunit sairaudet ja ylipaino.
Samalla on tärkeä huomauttaa noin kolmasosan tai puolien vaikeaa ja fataalia infektiomuotoa sairastaneista olleen aiemmin terveitä nuoria ja keski-ikäisiä.
Sanomattakin on selvää, että raskaana olevien komplikaatioriski nousee. Tälläinen riskien lisääntyminen nostaa myös sellaisen viruksen merkitystä, mikä ensisijaisesti infektoi nuoria ikäluokkia.
Lopuksi vaikka ei mitenkään pienimpänä huolena on se, ettei tiedetä edeltä, miten virus käyttäytyy kehitysmaissa esiintyvissä olosuhteissa. Tällä hetkellä suurin osa tapauksista on havaittu ja tutkittu eri maista jokseenkin hyvien olosuhteiden vallitessa.
Sallinette minun painottaa kahta seikkaa näistä monista. Ensinnäkin 99 % kaikesta äitikuolleisuudesta ilmenee kehitysmaissa, mikä on merkitsijä kehnosta äitiyshuollosta ja synnytysten hoidosta. Toiseksi noin 85% kroonisten tautien rasituksesta keskittyy matalan ja keskitason tuloja omaaviin maihin.
Vaikkakin pandemialla vaikuttaa olevan vain kohtalainen vaikeusaste näissä suhteellisen hyvin toimeentulevissa maissa, on järkevää olettaa ankeampaa kuvaa viruksen levittyä niille alueille, missä resursseja on rajoitetusti, terveyspolitiikka kehnompaa ja kaikenlaista taustalla olevaa lääketieteellistä ongelmatiikka on riittämiin.
Herrat ja Rouvat,
Eräs pandemioille luonteenomainen piirre on niitten nopea leviäminen maailman kaikkiin osiin. Viime vuosisadalla tämä leviäminen tapasi viedä 6-9 kuukautta silloinkin, kun kaikki kansainvälinen liikenne käytti laivoja ja junia.
Maitten tulisi varustautua havaitsemaan influenssatapauksia tai tulisi valmistua niitten esiintymisiin lähiaikoina. Niiden maitten, joissa influenssa-aalto on jo huippunsa ohittanut, pitäisi valmistautua toisen infketioaallon tulemiseen.
Kaikkien maiden terveysministeriöihin on toimitettu ohjeita spesifisistä suoja- ja varmuustoimenpiteistä. Varuillaan oloa odotetaan niiltä mailta, joissa ei vielä ole influenssatapauksia tai ehkä vain jokin harva tapaus.
Laaja-alaisia tartuntoja esiintyessä valtioitten pitäisi keskittyä potilaittensa asianmukaiseen hoitoon. Potilaiden testaamisia ja tutkimisia pitäisi prioritoida intensiiviresursseiksi välttäen joutumatta resurssipulaan kapasiteettien nopeasti revähdettyä.
WHO on keskustellut tiiviisti influenssarokotteen valmistajien kanssa. Ymmärtääkseni kausiluontoisen influenssan rokotteen tuotanto alkaa olla kohta jo tehty valmiiksi ja täydellä kapasiteetilla voidaan sitten varmistaa suurin mahdollinen pandemisen rokotteen toimitus tulevina kuukausina.
Rokotteiden saatavuuden ohella on myös useita ei –lääkkeellisiä interventioita ,jotka voivat suoda jotain suojaa virusta vastaan.
WHO ei edelleenkään aseta matkustamisille mitään rajoitussuosituksia eikä rajojen sulkemisia.
Influenssapandemiat olivat ne sitten kohtalaisia tai vakavia ovat merkittäviä tapahtumia, koska koko ihmiskuntaon miltei maailmanlaajuisesti infektioaltista.
Olemme kaikki samassa veneessä ja yhdessä tulemme tästä selviämäänkin.
Kiitos."
12.6.2006
WHO:n sanoma 11.kesäkuuta
Suomennosta: LÄHDE: Statement to the press by WHO Director-General Dr Margaret Chan
11 June 2009 World now at the start of 2009 influenza pandemic
"Hyvät Naiset ja Herrat,
Huhtikuun lopussa WHO julisti hälytystilan uuden A influenssaviruksen takia.Tämä erityinen H1N1 kanta ei ole aiemmin kiertänyt ihmiskuntaa. Virus on aivan uusi. Virus on tarttuvainen ja leviää helposti ihmisestä toiseen ja maasta toiseen. Tänään on noin 30 000 varmistettua tapausta raportoitu yhteensä 74 eri maasta. Tässä on vain osa asiaa. Muutamaa poikkeusta huomioonottamatta niissä maissa, joissa on esiintynyt suuri määrä sairaustapauksia tilanteen valvonta ja näytteitten otto on hyvä. Leviämistä maasta toiseen ei enää voida jäljittää selkeästi ihmisten tartuntaketjua määrittämällä. Jatkoleviäminen on katsottava väistämättömäksi tapahtumaksi. Olen neuvotellut johtavien influenssa-asiantuntijoiden, virologien ja kansanterveysviranomaisten kanssa. Kansainvälisten terveyttä koskevien säädösten (International Health Regulations) käynnistämien proseduurien mukaisesti olen konsultoinut Poikkeustilakomiteaa (Emergency Committee) , joka on tätä tarkoitusta varten perustettu. Saatavilla olevan näytön ja niihin tutustuneitten asiantunijoitten en arvion perusteella on influenssapandemian tieteelliset kriteerit täyttyneet
Olen sen takia päättänyt kohottaa influenssapandemian hälytystasolta (5) pandemiatasoon (6). Nyt 11. kesäkuuta 2009 maailma on vuoden 2009 influenssapandemiansa alussa. (START).
Olemme nyt pandemian varhaispäivissä Virus leviää tiukan ja huolellisen kontrollin alaisena. EI KOSKAAN ole havaittu pandemiaa näin varhain eikä näin tarkasti seurattu juuri tapahtuvalla ajalla aivan varhaisista alkuhetkistä alkaen. Nyt maailmassa saadaan niittää pandemiavalmiuden etuja, joihin ollaan investoitu viime kuluneen viiden vuoden aikana. Meillä on etumatkaa. Tämä antaa meille vahvan aseman. Mutta samalla se haastaa meitä tietojemme ollessa rajallisia ja tieteellisen epävarmuuden vallitessa saamaan lisää neuvoja ja uusia vakuuksia
Hyvän monitoroinnin , perusteellisten tutkimusten ja eri maitten suoran raportoinnin ansiosta olemme saaneet muutamia varhaisia pikakäsityksiä viruksen leviämisestä ja sen mahdollisesti aiheuttamasta tautikirjosta.
Tiedämme myös , että varhaisen siellä täällä esiintyvän viruksen tavat voivat muuttua äkisti. Virus tekee säännöt ja tämäkin H1N1 kaikkien influenssavirusten tapaan voi muuttaa sääntöjään sen suuremmitta sävelittä milloin tahansa.
Globaalisti ottaen meillä on hyvät syymme uskoa tämän pandemian ainakin varhaispäivinään pysyttelevän asteeltaan kohtalaisen vakavana. Kokemuksesta tiedämme vakavuusasteen vaihtelevan maasta toiseen monista tekijöistä johtuen. Nykytietämyksen mukaan potilaitten valtaenemmistöllä on ollut lieviä oireita ja toipuminen on ollut nopeaa ja täydellistä usein ilman mitään lääkehoitoakin.
Kautta mailman on menehtyneitten luku ollut pieni. Joka ainoa näistä kuolemista on kuitenkin traaginen ja meidän on koottava voimia kohdatessamme niitä yhä lisää. Kuitenkaan emme odota mitään vaikeiden tai fataalien infektioiden äkillistä ja kohtalokasta lisääntymistä.
Tiedämme uuden H1N1 viruksen infektoivan nuorta väkeä. Miltei kaikilla laajemman ja pitkäaikaisemman purkauksen alueilla pääosa infektoituneesta väestöstä on ollut alle 25 vuotiaita.
Joissain maissa noin 2% infektoituneista on saanut vakavamman taudin, jossa esiintyy nopea paheneminen kohti henkeä uhkaavaa keuhkokuumetta.
Useimmat vaikeat tai fataalit infektiot ovat kohdanneet aikuisia, jotka ovat 30-50 vuoden ikäisiä.
Tällainen tautimalli on merkitsevästi kausiluontoisesta influenssaepidemiasta poikkeava, missä useimmat menehtymiset kohtaavat iäkästä väestöä.
Monesti, tosin ei jokaisessa tapauksessa, vakava sairastuminen on tapahtunut siinä väestössä, jolla on jokin krooninen perussairaus . Alustaviin rajallisiin tietoihin pohjautuen voidaan sanoa, että näihin sairauksiin kuuluu hengitysteiden taudit, varsinkin astma, sydän-ja verenkiertoelinten taudit, diabetes, autoimmuunit sairaudet ja ylipaino.
Samalla on tärkeä huomauttaa noin kolmasosan tai puolien vaikeaa ja fataalia infektiomuotoa sairastaneista olleen aiemmin terveitä nuoria ja keski-ikäisiä.
Sanomattakin on selvää, että raskaana olevien komplikaatioriski nousee. Tälläinen riskien lisääntyminen nostaa myös sellaisen viruksen merkitystä, mikä ensisijaisesti infektoi nuoria ikäluokkia.
Lopuksi vaikka ei mitenkään pienimpänä huolena on se, ettei tiedetä edeltä, miten virus käyttäytyy kehitysmaissa esiintyvissä olosuhteissa. Tällä hetkellä suurin osa tapauksista on havaittu ja tutkittu eri maista jokseenkin hyvien olosuhteiden vallitessa.
Sallinette minun painottaa kahta seikkaa näistä monista. Ensinnäkin 99 % kaikesta äitikuolleisuudesta ilmenee kehitysmaissa, mikä on merkitsijä kehnosta äitiyshuollosta ja synnytysten hoidosta. Toiseksi noin 85% kroonisten tautien rasituksesta keskittyy matalan ja keskitason tuloja omaaviin maihin.
Vaikkakin pandemialla vaikuttaa olevan vain kohtalainen vaikeusaste näissä suhteellisen hyvin toimeentulevissa maissa, on järkevää olettaa ankeampaa kuvaa viruksen levittyä niille alueille, missä resursseja on rajoitetusti, terveyspolitiikka kehnompaa ja kaikenlaista taustalla olevaa lääketieteellistä ongelmatiikka on riittämiin.
Herrat ja Rouvat,
Eräs pandemioille luonteenomainen piirre on niitten nopea leviäminen maailman kaikkiin osiin. Viime vuosisadalla tämä leviäminen tapasi viedä 6-9 kuukautta silloinkin, kun kaikki kansainvälinen liikenne käytti laivoja ja junia.
Maitten tulisi varustautua havaitsemaan influenssatapauksia tai tulisi valmistua niitten esiintymisiin lähiaikoina. Niiden maitten, joissa influenssa-aalto on jo huippunsa ohittanut, pitäisi valmistautua toisen infketioaallon tulemiseen.
Kaikkien maiden terveysministeriöihin on toimitettu ohjeita spesifisistä suoja- ja varmuustoimenpiteistä. Varuillaan oloa odotetaan niiltä mailta, joissa ei vielä ole influenssatapauksia tai ehkä vain jokin harva tapaus.
Laaja-alaisia tartuntoja esiintyessä valtioitten pitäisi keskittyä potilaittensa asianmukaiseen hoitoon. Potilaiden testaamisia ja tutkimisia pitäisi prioritoida intensiiviresursseiksi välttäen joutumatta resurssipulaan kapasiteettien nopeasti revähdettyä.
WHO on keskustellut tiiviisti influenssarokotteen valmistajien kanssa. Ymmärtääkseni kausiluontoisen influenssan rokotteen tuotanto alkaa olla kohta jo tehty valmiiksi ja täydellä kapasiteetilla voidaan sitten varmistaa suurin mahdollinen pandemisen rokotteen toimitus tulevina kuukausina.
Rokotteiden saatavuuden ohella on myös useita ei –lääkkeellisiä interventioita ,jotka voivat suoda jotain suojaa virusta vastaan.
WHO ei edelleenkään aseta matkustamisille mitään rajoitussuosituksia eikä rajojen sulkemisia.
Influenssapandemiat olivat ne sitten kohtalaisia tai vakavia ovat merkittäviä tapahtumia, koska koko ihmiskuntaon miltei maailmanlaajuisesti infektioaltista.
Olemme kaikki samassa veneessä ja yhdessä tulemme tästä selviämäänkin.
Kiitos."
torsdag 11 juni 2009
WHO arvioi pandemisen viruksen vaarallisuutta
Suomennosta WHO:n artikkelista, jossa arvioidaan pandemisen influenssan vakavuutta. Assessing the severity of an influenza pandemic 11 May 2009IFLUENSSAPANDEMIAN VAKAVUUDEN ARVIOINNISTA 11 toukokuutta 2009
Properties of the virus VIRUKSEN OMINAISUUKSISTA
Population vulnerability VÄESTÖN HAAVOITTUVUUS
Subsequent waves of spread PANDEMIAN TOISTUVAT AALLOT
Capacity to respond KYKY VASTATA PANDEMIAN HAASTEESEEN
Assessment of the current situation. TÄMÄN HETKEN TILANNE
(1) The major determinant of the severity of an influenza pandemic, as measured by the number of cases of severe illness and deaths it causes, is the inherent virulence of the virus. However, many other factors influence the overall severity of a pandemic’s impact.
Päämerkitsijä pandemisen influenssan vakavuudesta vaikeiden tautitapausten ja menehtyneitten lukumääränä ilmaistuna kuvaa virukselle ominaista virulenssia.
(2) Even a pandemic virus that initially causes mild symptoms in otherwise healthy people can be disruptive, especially under the conditions of today’s highly mobile and closely interdependent societies. Moreover, the same virus that causes mild illness in one country can result in much higher morbidity and mortality in another. In addition, the inherent virulence of the virus can change over time as the pandemic goes through subsequent waves of national and international spread.
Sellainenkin pandeminen virus, joka vaikuttaa alun alkaen lieväoireiseltä muuten terveillä, saattaa olla yhteiskuntaa häiritsevä varsinkin aikamme hyvin liikkuvaisessa ja keskinäisistä kommunikaatioista riippuvaisessa yhteiskunnassa. Sitäpaitsi sama virus voi yhdessä maassa aiheuttaa lievää tautia ja toisessa paljon enemmän sairastuvuutta (morbiditeettiä) ja kuolleisuutta (mortaliteettia) .
Properties of the virus VIRUKSEN OMINAISUUKSISTA
(3) An influenza pandemic is caused by a virus that is either entirely new or has not circulated recently and widely in the human population. This creates an almost universal vulnerability to infection. While not all people ever become infected during a pandemic, nearly all people are susceptible to infection.
Influenssa on pandeminen, jos sen aiheuttajavirus on joko aivan uusi virus tai sitä ei ole äskettäin ollut kiertämässä kautta ihmiskunnan. Tällainen tilanne luo miltei globaalin alttiuden ihmisten infektoitumiselle. Toisaalta ei koskaan joka-ainut ihminen sairastu pandemiankaan aikana, mutta kuitenkin miltei kaikki ovat alttiita (vastaanottavaisia) saaman infektion.
(4) The occurrence of large numbers of people falling ill at or around the same time is one reason why pandemics are socially and economically disruptive, with a potential to temporarily overburden health services.
Koska pandemioissa ihmisiä sairastuu samaan aikaan suurin joukoin, pandemiat ovat yhteiskuntaa sosioekonomisesti häiritseviä ja hetkittäin saattavat aiheuttavaa terveydenhoidollisiin palveluihin ylirasituksen.
(5) The contagiousness of the virus also influences the severity of a pandemic’s impact, as it can increase the number of people falling ill and needing care within a short timeframe in a given geographical area. On the positive side, not all parts of the world, or all parts of a country, are affected at the same time.
Viruksen tarttuvuus vaikuttaa myös pandemiassa, koska silloin voi lyhyessäkin ajassa jollain geografisella alueella sairastuvien ja hoitoa tarvitsevien määrä lisääntyä tuntuvasti. Mutta on hyvä tietää, että ei joka puolella maailmaa eikä yhden maan joka osissa kaikki sairastu yhtä aikaa.
(6) The contagiousness of the virus will influence the speed of spread, both within countries and internationally. This, too, can influence severity, as very rapid spread can undermine the capacity of governments and health services to cope.
Viruksen tarttuvaisuus vaikuttaa sen leviämisnopeuteen valtioitten sisällä ja kansainvälisesti. Sekin vaikuttaa taudin vaikeusasteeseen, koska hyvin nopea leviäminen heikentää hallitusten ja terveyapalveluitten mahdollisuuksia pysyä ajan tasalla.
(7) Pandemics usually have a concentrated adverse impact in specific age groups. Concentrated illnesses and deaths in a young, economically productive age group will be more disruptive to societies and economies than when the very young or very old are most severely affected, as seen during epidemics of seasonal influenza.
Pandemioissa on tyypillistä, että ne keskittyvät jonkin tietyn ikäryhmän kimppuun. Nuoren, ekonomisesti tuottavan ikäryhmän sairastumiset ja menehtymiset heikentävät yhteiskuntia ja taloutta enemmän. Tavalliset kausiluontoiset influenssaepidemiat tapaavat taas kohdata hyvin pieniä lapsia ja hyvin iäkkäitä henkilöitä vakavammin.
Population vulnerability VÄESTÖN HAAVOITTUVUUS
(8) The overall vulnerability of the population can play a major role. For example, people with underlying chronic conditions, such as cardiovascular disease, hypertension, asthma, diabetes, rheumatoid arthritis, and several others, are more likely to experience severe or lethal infections. The prevalence of these conditions, combined with other factors such as nutritional status, can influence the severity of a pandemic in a significant way.
Väestön yleinen haavoittuvuus (sairastumisalttius) saattaa myös olla suurena tekijänä. Esimerkiksi on todennäköisempää, että sairastutaan vakavasti tai menehdytään infektioon, jos on jokin krooninen sairaus taustalla kuten sydän-ja verenkiertoelinten tauti, korkea verenpaine, astma, diabetes, nivelreuma tai moni muu tauti. Tällaisten sairaustilat liittyneenä muihin tekijöihin kuten ravitsemukselliseen tilaan voi vaikuttaa pandemian vakavuusasteeseen merkitsevästi.
Subsequent waves of spread PANDEMIAN TOISTUVAT AALLOT
(9) The overall severity of a pandemic is further influenced by the tendency of pandemics to encircle the globe in at least two, sometimes three, waves. For many reasons, the severity of subsequent waves can differ dramatically in some or even most countries.
Pandemian vakavuuteen kaiken kaikkiaan vaikuttaa niitten pyrkimys kiertää maapallo ainakin parina kolmena aaltona. Joissain tai useimmissa maissa näitten peräkkäisten pandemia-aaltojen vakavuus voi vaihdella dramaattisesti monesta syystä.
(10) A distinctive feature of influenza viruses is that mutations occur frequently and unpredictably in the eight gene segments, and especially in the haemagglutinin gene. The emergence of an inherently more virulent virus during the course of a pandemic can never be ruled out.
Influenssaviruksille tyypillinen piirre on niitten kahdeksassa geenisegmentissä usein tapahtuvat ja ennalta arvaamattomat mutaatiot . Tämä pätee varsinkin hemagglutiniinigeeniin.
(11) Different patterns of spread can also influence the severity of subsequent waves. For example, if schoolchildren are mainly affected in the first wave, the elderly can bear the brunt of illness during the second wave, with higher mortality seen because of the greater vulnerability of elderly people.
Toistuvilla pandemia-aalloilla esiintyvät erilaiset leviämistavat voivat myös vaikuttaa pandemian vakavuuteen. Jos esimerkiksi enismmäinen aalto on kohdannut koululaisia, voi toisessa aallon isku kohdistua iäkkäisiin, jolloin heillä saattaa olla suurempi mortaliteetti, koska iäkkäämmät ovat haavoittuvampia.
(12) During the previous century, the 1918 pandemic began mild and returned, within six months, in a much more lethal form. The pandemic that began in 1957 started mild, and returned in a somewhat more severe form, though significantly less devastating than seen in 1918. The 1968 pandemic began relatively mild, with sporadic cases prior to the first wave, and remained mild in its second wave in most, but not all, countries.
Viime vuosisadalla vuoden 1918 pandemia alkoi lievänä ja palasi kuuden kuukauden kuluttua paljon tuhoavampana. Vuoden 1957 pandemia alkoi lievänä ja palasi jonkin verran vaikeampana, tosin vuoden 1918 pandemiaan verraten paljon vähemmän tuhoavana. Vuoden 1968 pandemiakin alkoi lievänä ja sitä edelsi jotkut sporadit tapaukset ennen ensimmäistä aaltoa ja se pysyteli lievänä useimmisa maissa, ei tosin kaikissa.
Capacity to respond KYKY VASTATA PANDEMIAN HAASTEESEEN
(13) Finally, the quality of health services influences the impact of any pandemic. The same virus that causes only mild symptoms in countries with strong health systems can be devastating in other countries where health systems are weak, supplies of medicines, including antibiotics, are limited or frequently interrupted, and hospitals are crowded, poorly equipped, and under-staffed.
Terveyspalvelujen laatu on se, mikä vaikuttaa pandemian yllättäessä. Maissa, joissa terveydenhoidolliset järjestelmät toimivat hyvin, sama virus voi aiheuttaa lieviä oireita ja päinvastaisesa tilanteessa virus voi olla tuhoisa lääkkeitten ja antibioottien saannin ollessa rajoitettua tai keskeytynyt, sairaaloiten ollessa täpötäynnä, huonosti varustettuja ja henkilökuntaapuuttuu.
Assessment of the current situation. TÄMÄN HETKEN TILANNE
(14) To date, the following observations can be made, specifically about the H1N1 virus, and more generally about the vulnerability of the world population. Observations specific to H1N1 are preliminary, based on limited data in only a few countries.
Seuraavaa voidaan havaita erityisesti viruksesta H1N1 ja maailman väestön haavoittuvuudesta. Virusta H1N1 koskevat obeservaatiot ovat preliminäärisiä ja perustuvat muutamien maiten tietoihin.
(15) The H1N1 virus strain causing the current outbreaks is a new virus that has not been seen previously in either humans or animals. Although firm conclusions cannot be reached at present, scientists anticipate that pre-existing immunity to the virus will be low or non-existent, or largely confined to older population groups.
Viruskanta H1N1, joka aiheuttaa nykyista purkausta on uusi virus, jota ei ole aiemmin tavattu niin ihmisistä kuin eläimistäkään. Vaikka varmoja johtopäätöksiä ei voi ( 9.5. 2009) tehdä, tiedemiehet aavistelevat, että olemassaolevaa immuniteettiä virusta kohtaan ei ole tai että se on vain matalaa tai suurimmaksi osaksi rajoittuu iäkkäämpään väestöön.
(16) H1N1 appears to be more contagious than seasonal influenza. The secondary attack rate of seasonal influenza ranges from 5% to 15%. Current estimates of the secondary attack rate of H1N1 range from 22% to 33%.
Virus H1N1 näyttää olevan tarttuvampi kuin tavallinen kausiluontoinen influenssavirus. Kausiluontoisen influenssan toisen hyökkäysaallon tahti on 5-15%. Tämänhetkisten arvioitten mukaan toinen hyökkäystähti tämän H1N1 viruksen taholta tulee olemaan 22- 33%.
(17) With the exception of the outbreak in Mexico, which is still not fully understood, the H1N1 virus tends to cause very mild illness in otherwise healthy people. Outside Mexico, nearly all cases of illness, and all deaths, have been detected in people with underlying chronic conditions
Mexikossa tapahtunutta taudinpurkausta lukuunottamatta (mitä muuten ei ole täysin vielä pystytty käsittämään) H1N1 virus tapaa aiheuttaa hyvin lievän taudin muuten terveissä ihmisissä. Mexikon ulkopuolella melkein kaikki sairastumiset ja kuolemantapaukset ovat kohdanneet sellaisia henkilöitä, jolla on jokin krooninen tauti.
(18) In the two largest and best documented outbreaks to date, in Mexico and the United States of America, a younger age group has been affected than seen during seasonal epidemics of influenza. Though cases have been confirmed in all age groups, from infants to the elderly, the youth of patients with severe or lethal infections is a striking feature of these early outbreaks.
Kahdessa tämän hetken suurimassa ja parhaiten dokumentoidussa taudinpurkauksessa, Mexikon ja USA:n influenssassa, on sairastunut nuorempaa ikäluokkaa kuin kausiluontoisessa influenssassa tapaa sairastua. Vaikka on tavattu sairaustapauksia kaikista ikäluokista vauvoista iäkkäisiin, niin vakavaan tai letaaliin infektion sairastuneitten potilaiden nuori ikä on silmiinpistävä piirre näissä varhaisissa purkauksissa.
(19) In terms of population vulnerability, the tendency of the H1N1 virus to cause more severe and lethal infections in people with underlying conditions is of particular concern.
Väestön haavoittuvuus huomioonottaen on syytä panna merkille H1N1 viruksen pyrkimys aihuttaa vakavampia ja letaaleja infektioita niille, joilla on taustasairaus .
(20) For several reasons, the prevalence of chronic diseases has risen dramatically since 1968, when the last pandemic of the previous century occurred. The geographical distribution of these diseases, once considered the close companions of affluent societies, has likewise shifted dramatically. Today, WHO estimates that 85% of the burden of chronic diseases is now concentrated in low- and middle-income countries. In these countries, chronic diseases show an earlier average age of onset than seen in more affluent parts of the world.
Useista syistä kroonisten tautien prevalenssi on noussut dramaattisesti sitten vuoden 1968, jolloin edellisen vuosisadan viimeinen pandemia pyyhki yli maapallon. Näiden aiemmin hyvinvointisairauksina pidettyjen tautien geografinen jakauma on myöskin vaihtunut huomattavasti. Nykyään WHO arvioi, että 85% kroonisten tautien rasitteesta keskittyy matala-ja keskituloisten kansojen maihin. Niissä maissa krooniset taudit alkavat keskimäärin varhemmalla iällä kuin paremminvoivissa maailman osissa.
(21) In these early days of the outbreaks, some scientists speculate that the full clinical spectrum of disease caused by H1N1 will not become apparent until the virus is more widespread. This, too, could alter the current disease picture, which is overwhelmingly mild outside Mexico.
Näinä taudinpurkauksen varhaisina päivina (9. 5. 2009) jotkut tiedemiehet spekuloivat H1N1 viruksen aiheuttaman taudin täyden kliinisen kirjon ilmenevän vasta, kun virus on laajemmalle levinnyt. Silloin voi taudin nykyinen kuva muuntua, nythän se on Meksikon ulkopuolella hämmästyttävän lievä.
(22) Apart from the intrinsic mutability of influenza viruses, other factors could alter the severity of current disease patterns, though in completely unknowable ways, if the virus continues to spread.
Muutkin tekijät kuin influenssaviruksen sisäinen mutatoituvuus voivat muuntaa tämän hetken taudinkuvan vaikeusastetta, vieläpä täysin tuntemattomilla tavoilla, jos virus jatkaa leviämistään.
(23) Scientists are concerned about possible changes that could take place as the virus spreads to the southern hemisphere and encounters currently circulating human viruses as the normal influenza season in that hemisphere begins.
Tiedemiehet ovat ottaneet huomioon mahdolliset muutokset, joita voit tapahtua, kun virus leviää eteläiselle pallonpuoliskolle kattaen sikäläisen tavallisen kausiluontoisen influenssan alueita.
(24) The fact that the H5N1 avian influenza virus is firmly established in poultry in some parts of the world is another cause for concern. No one can predict how the H5N1 virus will behave under the pressure of a pandemic. At present, H5N1 is an animal virus that does not spread easily to humans and only very rarely transmits directly from one person to another.
Lintuinfluenssa H5N1 on toinen seikka, mikä on otettava huomioon. Sitä on pinttyneenä siipikarjassa eräin paikoin maailmaa. Ei ole ketään, joka pystyy ennalta sanomaan, miten H5N1 tulee käyttäytymään pandemisessa paineessa. Tällä hetkellä H5N1 on zoonoottinen virus( eläin virus) , joka ei leviä ihmiseen helposti; vain hyvin harvoin se välittyy suoraan henkilöstä toiseen.
Prenumerera på:
Kommentarer (Atom)