GOLGIn laite ja SARS 2 interaktioproteiinit SARS2
interaktioproteiineissa mainitaan seuraavat interaktioproteiinit:
Giantin, GCP372, GOLGB1 , golgin B1:
(Sars2 nsp3)
GRASP65, GORASP1, GOLGI
Reassembly Stacking protein 11. (Sars2 nsp13)
Golgin -95, Golgin A2, GM130, GOLGA2,
(Sars2 nsp13)
GOLGA7, Golgin subfamily A member 7. Golgin A7
(Sars2 Spike)
Golgin-160, GOLGA3, Golgin A3
(Sars2 nsp13)
GCC2,( GRIP and
coiled coil domain containing 2, GCC185, GOLGI Coiled Coil protein
185( interaktioproteiini: Sars 2 CoV nsp 13
GCC1 , (GCC88,
GOLGI Coiled coil protein 88 ) (Sars2 nsp 13)
https://www.genecards.org/cgi-bin/carddisp.pl?gene=GCC1&keywords=GCC88
(Sars2 nsp13 interaktioproteiini)
WASH complex
subunit 4 (Sars2 nsp 2)
(GOLG4 ja TMF sitovat tekijöitä, jotka sitoutuvat WASH kompleksiin. )
Muita :
GMAP-210, TRIP-11, GOLGI- microtubule associated protein 210 kDa.
GOLGIN A4, Golgin -245, GOLGA4. https://www.genecards.org/cgi-bin/carddisp.pl?gene=GOLGA4&keywords=GOLGA4
Golgin-84, Golgin5, GOLGA5, RETII, Ret fused gene 5 protein, https://www.genecards.org/cgi-bin/carddisp.pl?gene=GOLGA5&keywords=GOLGA5
Review ARTICLE
Front. Cell Dev. Biol., 18 June 2019 |
https://doi.org/10.3389/fcell.2019.00094
The Physiological Functions of the Golgin Vesicle Tethering Proteins Martin Lowe*
- Faculty of Biology, Medicine and Health, University of
Manchester, Manchester, United Kingdom
{kind=link}
Tethering Proteins
Martin Lowe*- Faculty of Biology, Medicine and Health, University of
Manchester, Manchester, United Kingdom
The Golgi apparatus lies at the heart of the secretory pathway, serving to modify newly synthesized cargo proteins and to sort and transport these proteins to their final destination, which may be inside or outside the cell. It is comprised of flattened membrane compartments called cisternae that are layered on top of one another to form the Golgi stack. In non-vertebrates the Golgi stacks exist as discrete entities within the cytoplasm, whereas in vertebrates the stacks are laterally connected to form a single-copy Golgi ribbon which is located adjacent to the centrosome (Figure 1). Newly synthesized cargo proteins arriving from the endoplasmic reticulum (ER) enter the Golgi apparatus at the cis-face, which in vertebrates comprises a tubulo-vesicular compartment called the cis-Golgi network (CGN). Cargo then transits the Golgi stack before arriving at the exit station of the Golgi, the trans-Golgi network (TGN), where it is sorted into carriers for delivery to its post-Golgi destination. As cargo transits the Golgi stack, numerous resident enzymes carry...
Golgins as Vesicle Tethering Proteins
https://www.frontiersin.org/files/Articles/459614/fcell-07-00094-HTML/image_m/fcell-07-00094-g002.jpg
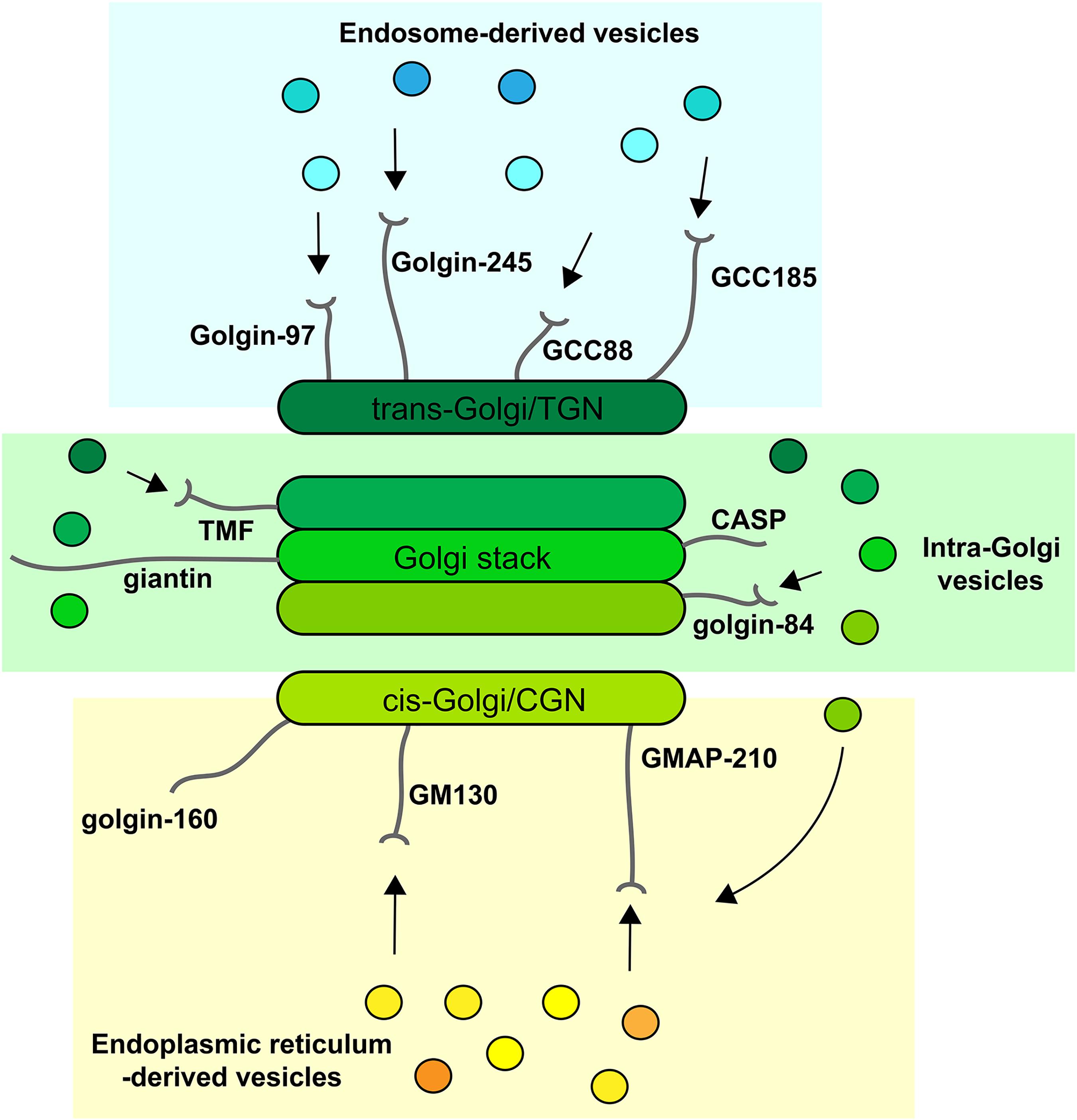
The golgins comprise a family of Golgi-localized coiled-coil proteins with a similar topology (Munro, 2011; Witkos and Lowe, 2015).
They are anchored to the Golgi membrane by their carboxy-terminus, and
extend into the cytoplasm to facilitate vesicle capture, which is in
most cases is mediated by the extreme amino-terminus of the protein (Cheung et al., 2015; Gillingham and Munro, 2016; Wong et al., 2017; Gillingham, 2018).
In humans, there are at least 11 golgins, with varying degrees of
conservation between different eukaryotes depending upon the particular
golgin. The different golgins are localized to distinct regions of the
Golgi apparatus, consistent with their ability to tether different
vesicle types (Wong and Munro, 2014; Gillingham and Munro, 2016; Gillingham, 2018). For example, golgins localized at the cis-Golgi
are competent to selectively tether vesicles arriving from the ER and
intra-Golgi vesicles mediating recycling from later Golgi cisternae,
whereas those at the trans-Golgi tether vesicles arriving from the endolysosomal system (Figure 2).
In contrast, golgins residing within the Golgi stack are able to tether
only intra-Golgi transport vesicles. Golgins therefore play a major
role in dictating the specificity of vesicle traffic within the
secretory pathway. In addition, the elongated nature of the golgins,
coupled with their ability to tether vesicles via their membrane-distal
amino-termini, giving a greater radius of capture, allows for increased
efficiency of traffic. Following vesicle capture, golgins are thought to
cooperate with other proteins, including Rab GTPases and multi-subunit
tethering complexes such as COG and GARP, to mediate the transition from
tethering to membrane fusion, which operates over a relatively short
distance and is mediated by SNARE proteins (Witkos and Lowe, 2017).
Inga kommentarer:
Skicka en kommentar