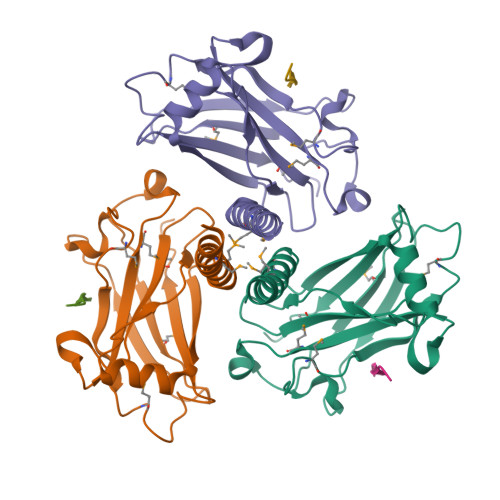
https://www.ebi.ac.uk/interpro/entry/InterPro/IPR032500/
hort name: bCoV_S1_N
Overlapping homologous superfamilies
domain relationships
Description
This
entry represents the N-terminal domain of the betacoronavirus-like
trimeric spike glycoprotein. The distal S1 subunit of the coronavirus
spike protein is responsible for receptor binding. S1 contains two
domains; an N-terminal galectin-like domain (NTD) and a receptor-binding
domain (S1 RBD) also referred to as the S1 CTD or domain B. Either the
S1 NTD or S1 RBD, or occasionally both, are involved in binding the host
receptors. S1 NTD is located on the side of the spike trimer and mainly
recognises sugar receptors
[1, 2, 3, 4].
For many betacoronaviruses (bCoVs), for example mouse hepatitis virus
(MHV), the RBD is located in the NTD. The structure of the MHV S1 NTD
showed the same fold as human galectins (galactose-binding lectin),
however it does not bind any sugar; instead, it binds to the
carcinoembryonic antigen cell-adhesion molecule (CEACAM1) through
protein-protein interactions
[2].
All three CEACAM21a-binding sites in MHV spikes can be fully occupied
by CEACAM1a. It has been shown that CEACAM1a binding to the MHV spike
weakens the interactions between S1 and S2 and facilitates the
proteolysis of the spike protein and dissociation of S1
[2].
The homologous bovine CoV (BCov) S1 NTD also possesses a galectin fold
but binds to sialic acid-containing moieties on host cell membranes, as
does the NTD of three other group A b-Covs, namely human CoV (HCoV)
OC43, avian b-CoV, and infectious bronchitis virus (IBV)
[5].
Despite the S1 NTD of human respiratory b-CoV HKU1 being highly
homologous to the NTDs of MHV and bovine CoV, it does not bind to either
sugar or human CEACAMs and the RBD is found instead in the S1 RBD
domain
[5]The
bCoV NTDs contain a conserved beta-sandwich core, but exhibit variant
folds in the peripheral elements located in the top-ceiling region and
on the lateral side. The core sandwich comprises in total sixteen
anti-parallel beta-strands, assembling into three (upper, middle, and
lower) beta-sheet layers. While showing different compositions and
structures, the peripheral elements are topologically equivalent
beta-sandwich-core insertions, highlighting a divergent evolution
process for bCoVs to form different lineages
[3].
Galektiini inhibiittorit, onko niistä sars-2 vastaisiksi lääkkeiksi?
file:///C:/Users/lea/AppData/Local/Temp/biomedicines-09-01208.pdf
Abstract: Galectin-3 is a carbohydrate-binding protein and the most studied member of the galectin
family. It regulates several functions throughout the body, among which are inflammation and post-
injury remodelling. Recent studies have highlighted the similarity between Galectin-3′s carbohydrate
recognition domain and the so-called “galectin fold” present on the N-terminal domain of the S1
sub-unit of the SARS-CoV-2 spike protein. Sialic acids binding to the N-terminal domain of the
Spike protein are known to be crucial for viral entry into humans, and the role of Galectin-3 as a
mediator of lung fibrosis has long been the object of study since its levels have been found to be
abnormally high in alveolar macrophages following lung injury. In this context, the discovery of
a double inhibitor may both prevent viral entry and reduce post-infection pulmonary fibrosis. In
this study, we use a database of 56 compounds, among which 37 have known experimental affinity
with Galectin-3. We carry out virtual screening of this database with respect to Galectin-3 and Spike
protein. Several ligands are found to exhibit promising binding affinity and interaction with the
Spike protein’s N-terminal domain as well as with Galectin-3. This finding strongly suggests that
existing Galectin-3 inhibitors possess dual-binding capabilities to disrupt Spike–ACE2 interactions.
Herein we identify the most promising inhibitors of Galectin-3 and Spike proteins, of which five
emerge as potential dual effective inhibitors. Our preliminary results warrant further in vitro and
in vivo testing of these putative inhibitors against SARS-CoV-2 with the hope of being able to halt
the spread of the virus in the future.
Keywords: Galectin-3; spike; dual inhibitors; COVID-19; docking
1.1. Spike: Main Features and Functions
The Spike protein (also called S-protein) component of SARS-CoV-2 is a central target
in the fight against COVID-19 since it is the primary target of antibodies that provide
immunity against the virus. The surfaces of coronaviruses are covered with these spikes,
giving them their distinctive crown-like appearance in electron micrographs. The spikes
initiate the process of infection, binding to receptors and then fusing with the cell membrane to release the viral genome inside. Many other enveloped viruses, including influenza hemagglutinin and the envelope glycoproteins of HIV-1 and Ebola, use similar spike-likeproteins to infect cells.
The spike protein is composed of three identical chains that together form a complex with a small domain inside the virus, a membrane-spanning segment,
and a large ectodomain that extends outward from the virus.
The S-protein exists in a meta stable prefusion conformation that undergoes a substantial structural rearrangement
to fuse the viral membrane with the host cell membrane.
This process is triggered when the S1 sub-unit binds to a host cell receptor. Receptor binding destabilises the prefusion trimer, resulting in the shedding of the S1 sub-unit and transition of the S2 sub-unit to a stable postfusion conformation.
To engage a host cell receptor, the receptor-binding domain (RBD)
of S1 undergoes hinge-like conformational movements that transiently hide or expose
the determinants of receptor binding. These two states are referred to as the “down” (or
“closed”) conformation and the “up” (or “open”) conformation (Figure 1), where down
corresponds to the receptor-inaccessible state and up corresponds to the receptor accessible state, which is presumed to be less stable [5].
The S1 sub-unit of the spike protein of SARS-CoV-2, critical for its entry into host
cells, can be subdivided into an NTD and a C-terminal domain (CTD) (Figure 2).
The
role of the CTD in viral entry has been well characterised in the literature, as it binds
angiotensin-converting-enzyme 2 (ACE2) receptors.
ACE2 is an enzyme that activates angiotensin, a peptide hormone involved in the
control of blood pressure. ACE2 is found on lung, heart, kidney, and intestinal cells, making these cells the primary targets for infection by the virus [5,6]. In addition, the spike is a glycoprotein: the ectodomain is covered with sugar chains that help to mask the virus from the immune system [6]. Watanabe et al. reported 22 N linked glycosylation sites on
Spike protein. In their study, across the 22 N-linked glycosylation sites, 52% were found to be fucosylated, while 15% of the glycans contained at least one sialic acid residue. Sialic acids, it should be noted, have been described as “eccentric” in that they bind several different pathogens and toxins. Owing to their location and abundant distribution, sialic acids participate in a wide variety of physiological and pathological processes. Sialic
acids are a common constituent of glycoproteins, glycolipids, and gangliosides. They
Biomedicines 2021, 9, 1208 3 of 27
decorate the terminal of sugar chains at the surface of cells or soluble proteins.
Sialic acids
linked to glycoproteins and gangliosides are recruited by a broad spectrum of viruses—
including coronaviruses—as receptors and/or attachment factors for cell entry. In many
viral infections (influenza, Ebola, SARSCoV, among others), glycan-mediated interactions are essential for the initial contact between the virus and the host [7–9]. The S1-NTD of SARS-CoV-2 has been shown to possess a sialic acid binding site highly similar to thatobserved in MERS-CoV [10,11].
In MERS-CoV, depletion of sialic acids inhibits cell entry
by the virus. As such, the interaction between S1 -NTD and host sialic acids may be critical
for SARS-CoV-2 cell entry as a means of stabilizing the interaction between the S1 -CTD
and ACE2
...
The Gal-3 inhibitors developed have ranged from
the natural binding lactose motif to synthetic derivatives, such as monogalactosides and
thiodigalactosides adorned with featuring non-carbohydrate structural elements such as
aryl triazoles and aromatic amides [15]. The Galectin-3 protein is linked to COVID-19
pathogenesis in particular in terms of its role in Cytokine Storm Syndrome (CSS) and as amediator of lung fibrosis [13].