Paul Spearman et Eric O.Fred . HIV Interactions with host Cell proteins. Sivuilla 125146 kerrotaan taas minulle uudesta käsitteestä:
Artikkeli on Llano Manuel et al. kirjoittama Virological and Cellular Roles of the Transcriptional Coactivator LEDGF/p75.
Kyse on kromatiiniin assosioituneesta soluproteiiniesta, jonka tiedetään osallistuvan transkriptionaaliseen säätelyyn, solun elossapysymiseen ja autoimmuniteettiin. Nimikirjainten ryväs LEDGF/p75 tulee sanoista Lens Epithelium-Derived Growth Factor p75 ja se kuuluu HDGF-perheeseen, joka taas on seitsenjäseninen Hepatoma-Derived Growth Factor. Yleensäkin GF tarkoittaa kasvutekijää, Growth Factor.
Proteiinilla on siis tehtäviä niin solun kuin viruksen hyväksi ja avainpiirteenä on sen kyky toimia molekulaarisena adaptorina ja tether-proteiinina, eräänlaisena liekana tai ankkuriköytenä kromatiinisäikeeseen. Tässä artikkelissaan tiedemiehet selventävät LEDGF/p75 ja LEDGF/p52 nimisten proteiinien yhä laajemmiksi ymmärrettyjä rooleja erilaisissa soluprosesseissa ja tautitiloissa.
- Miten ja milloin tämä proteiini keksittiin?
Vuosina 1998- 2003 neljässä eri tutkimuksessa- jotka olivat aivan eri tutkimuskentistä ( transkription säädön, solun elossapysymisen, autoimmuniteetin ja virologian aihepiireistä ) tutkijat havaitsijat toisistaan riippumatta erään denaturoidussa SDS-geelissä migroituvan polypeptidin kooltaan kDa 75. Alkulöytö tapahtui kun tehtiin mikrosekventioimista proteiinille, joka puhdistui PC4:n kanssa samanaikaisesti. PC4 on yleisen transkriptionaalisen koaktivaattorin positiivinen co-faktori 4.
Vuonna 2000 havaittiin sen osuus autoimmuniteetissa. Identifioitiin p75 , kun oltiin seulomassa cDNA-kirjastoa ihmisseerumilla, joka oli reaktiivinen tuma-autoantigeeniproteiinia DFS70 kohtaan (DFS70= dense fine speckled protein 70kDa). Tästä tutkimuslinjasta pääteltiin LEDGF/75:llä olevan osuutta autoimmuniteetissä. Muut tutkimukset lisäksi osoittivat sillä olevan osuutta solun elossapysymisessä ja apoptoosin estossa.
Vuonna 2003 Cherepanov et al. eristivät p75-proteiinin "polyproteiinina", joka immunosaostui HIV-1 integraasin(IN) kanssa kun integraasia esiintyi yliexpressoituna koe-olosuhteissa muissa kuin virusyhteyksissä.
Nyt näihin proteiineihin:
Nykyään tiedetään myös, että lentivirukset, retrovirussuku johon HIV-1 kuuluu, on kaapannut itselleen LEDGF/p75 proteiinin tarjoamat kromatiiniin kiinnittymiskapasiteetit viruselämänsä pakollista isäntäsolun kromosomiin integroitumista varten (2006)
http://www.pnas.org/content/107/7/2735/F1.large.jpg
{kind=link}
Riippuvuutta LEDGF/P75 proteiinin tarjoamasta liekaamisavusta on kädellisten (primate) lentiviruksen lisäksi myös kahdella muulla ryhmällä kynsiapinoitten ( ungulate) ja kissojen (feline) lentiviruksilla.
- Miten LEDGF geenit järjestyvät ja minkälaisia pleissivariantit ovat?
Molempia koodaa geeni PSIP1 (PC4- ja SFRS-interaktion tekevä proteiini 1) ja se sijaitse ihmisen kromosomissa 9 p22.3. Siinä on 15 exonia ja 14 intronia.
http://en.wikipedia.org/wiki/Chromosome_9_%28human%29
http://atlasgeneticsoncology.org/Genes/PSIP1ID405ch9q22.html
LEDGF/p52 ja LEDGF/75 pleissivariantit omaavat samanlaisen N-terminaalin 325 aminohapon osalta.
Beeta-barrel, helices.CR1, NLS, väli, AT hooks, väli, CR2, CR3.
Lyhyt p53 omaa c-terminaalin, jossa on 8 aminohappoa.
N-terminaalinen osa omaa kromatiiniin (DNA-säikeeseen) sitoutumisominaisuuksia.
http://www.nature.com/mt/journal/v18/n3/images/mt201036f1.gif
{kind=link}
Pitkä pleissivariantti p 75 omaa C-terminaalin jossa on 205 aminohappoa. Tässä on integraasille oma sitova domaani, IBD 340- 417, mikä on keskeinen proteiinin merkityksessä virologisesti ja solun kannalta ( 2004, 2005).
niiten mRNA ja vastaavaa proteiinia esiintyy yleisesti kehossa, p75 esiintyy runsaampana useimmissa kudoksissa, muta p53 mRNA on suhteellisesti runsaampana tietyisä kudoksissa kuten aivoissa, thymuksessa, testiksessä. Pleissautuminen saattaa säätyä kudostyyppisesti.
Lyhyttä pleissivarianttia on tunnistettu promyelosyyttisistä leukemiasoluista mRNA-analyyseistä.
- Kromatiiniin sitoutumisesta
Tumaan menon (nuclear import) määrää vastasyntyneessä proteiinissä oleva NLS signaali ( nuclear localization signal) aminohapoissa 148- 156.
( Proteiinin pitää syntyä sytoplasmakoneistossa ja siinä vaiheessa saada tuo osoitelappu NLS - ja tästä on enemmänkin tarinaa Wikipediassa: http://en.wikipedia.org/wiki/Nuclear_localization_sequence Ilman tuota osoitelappua ei mikään rehellinen proteiini pääse tumaan ja vaikutaakseen rehellisesltä on HIV-viruksen hankittava NLS-osoitelappu itselleen ja tietysti se kaappaa sen).
http://www.nature.com/nrmicro/journal/v5/n3/images/nrmicro1579-f4.jpg
{kind=link}
LEDGF proteiinien NLS kuuluu klassiseen baasiseen (SV40 large T antigen) perheeseen ja tumaan kuljetus vaatii funktionaalisen Ran-proteiinin, adaptoriproteiini importiini-alfan ja ja tumaan kuljetusreseptorin importiini- beta.
Jos NLS poistettaisiin tieteellisesti pitäisi vastasyntyneen LEDGF/p75 proteiinin jäädä sytoplasman puolelle, jolloin sen funktio jäisi puuttumaan.
Kuitenkin jakautuvissa soluissa mitoosissa voidaan olla NLS-tekijää vaillakin lokalisaatiossa. Proteiini on silloin kromatiiniin kietoutuneena niin tehokkaasti tumaplasman ja sytoplasman sisältöjen mitoottisen sekoittautumisen aikana, että stabiilisti ilmentyvät NLS-mutantit nähdään vain tiiviisti liittyneenä kromosomeihin. Kromatiiniin sitoutumista välittää osittain N-terminaalinen PWWP-domaani ( aminohapot 1-93). PWWP-domaanilla onkyllä homologiaa siihen domaaniin, johon mm integraasi liittyy (IBD), mutta IN ja muut proteiinit dissosioituvat irti kromatiinista mitoosissa siitä kohdasta.
LEDGF/p75 omaa Triton- resistentin kromatiiniin sitoutumisen.
Solusyklivaihe G1: p52 on tuman periferiassa ja p75 diffuusisti kautta tuman ( dense fine speckled pattern).
http://schoolworkhelper.net/wp-content/uploads/2010/11/cell_cycle.jpg
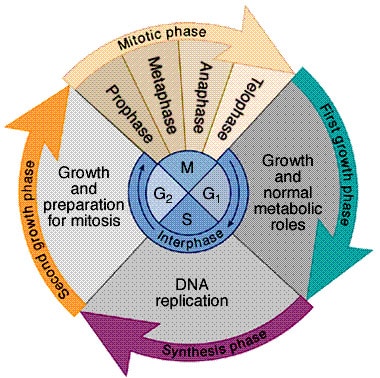{kind=link}
Metafaasi: P52 muodostaa sylinterimäisen mallin kromosomien ympärille ja p75 liittyy kromosomeihin juovallisesti.
http://en.wikipedia.org/wiki/Metaphase
Sytokineesi: P75 esiintyy diffuusina tumassa, mutta p52 säilyttää sylinterimäisen mallin.
http://upload.wikimedia.org/wikipedia/commons/thumb/e/e0/Cytokinesis_eukaryotic_mitosis.svg/800px-Cytokinesis_eukaryotic_mitosis.svg.png
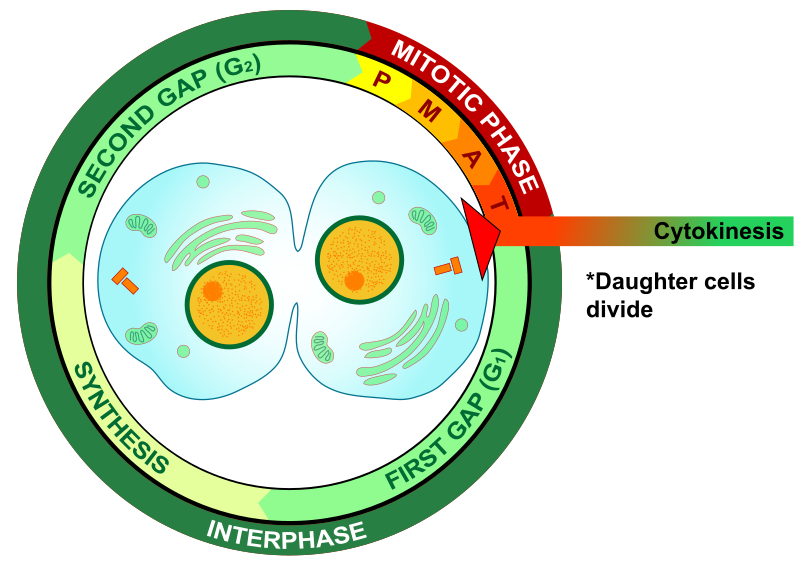{kind=link}
Kummankin C-terminaaliset alueet hallitsevat tuman sisäisen segrekoitumisen hienosäätelyjä.
P75 lokalisoituminen moduloituu muitten kromatiiniin sitoutuneitten proteiinien interaktioista TAI vaihtoehtoisesti p75-polyproteiinin pitkä C-terminaali voisi modifioida kummankin polyproteiinin N-terminaalien kesken jaettuja kromatiini-interaktioita. (Siis LEDGF-proteiineilla on funktionaalisesti kiinteä yhteistyö)
KUVA normaalista kromatiinista (DNA:n kärjestymisestä kromosomiksi)
http://themedicalbiochemistrypage.org/images/chromatin-structure.jpg
{kind=link}
Asian kliinisestä merkityksestä tässä:
http://www.mdpi.com/1999-4915/1/3/780/htm
- NYKYTILANNE tämän proteiinin uumoilta. Tuorein löytämäni PubMed hakutulos:
Source
Dipartimento Farmaco-Chimico, Università degli Studi di Messina, Italy. ldeluca@unime.it
Abstract
Although 25 compounds are currently licensed as anti-HIV drugs, the development of multidrug-resistant viruses, as well as their severe side effects, compromise their efficacy and limit treatment options. The search for new targets in order to cure AIDS has revealed that the inhibition of some protein-protein interactions in the HIV life cycle may provide an important new approach to fight this disease. The interaction between HIV-1 integrase (IN) and Lens Epithelium-Derived Growth Factor (LEDGF/p75) has increasingly gained attention as a valuable target for a novel anti-retroviral strategy. This article reviews the discovery and development of molecules capable of interrupting the LEDGF/p75-IN interaction reported to date.
Inga kommentarer:
Skicka en kommentar