Noroa norona NORO-viruksesta edelleen
NOROVIRUKSEN HOITOON SUUNNITELLAAN TERAPIAA.
NORO-viruksen tapoja ja sen vastaisen terapian hahmottelua käsittävä väitöskirja on antoisa ja koetan suomentaa siitä joitain kohtia, että saa käsitystä siitä, minkälaisen agenssin kanssa ollaan tekemisissä kun sanotaan että NORO-viruksesta on jälleen uutta uhkaa.
Väitöskirjan tekijä on Gustav E. Rydell et al. täällä Göteborgissa 2009.
Hänen väitöskirjansa nimi englanniksi on
Norovirus, causative agent of winter vomiting disease, exploits several histo-blood group glycans for adhesion.
Työ on tehty biomedisiinisessä instituutissa kliinisen kemian ja transfuusiolääketieteen osastolla Göteborgin Yliopistossa Sahlgrenskan Akatemiassa. Sahlgrenskan Keskuskirjastoon on kirja leimattu 5.11. 2009 sisään, joten asia on hyvin tuoretta.
Väitöskirjaa käsittää osatutkimukset, jotka tässä luettelen:
(I) Vuodelta 2006 Larsson MM, Rydell GEP et al.
Antibody prevalence and titer to norovirus (genotype II) correlate with secretor (FUT2) but not with ABO phenotype or Lewis (FUT3) genotype. The Journal of Infectious Diseases 194(10) pages 1422-7.Tästä olen aiemmin tehnyt lyhyttä mainintaa suomeksi.
(II) Vuodelta 2009: Rydell GE, Nilsson J et al.
Human noroviruses recognize sialyl Lewis x neoglycoprotein, Glycobiology 19(3), pages 309-20.
(III) Vuodelta 2009: Nilsson J, Rydell GE et al.
Norwalk-virus like particles bind specifically to A, H and difucosylated Lewis but not to B histo-blood group active glycosphingolipids. Glycoconjugate Journal, Apr 23 DOI 10.1007/s 10719-009-9237-x.
(IV) (2009) Rydell GE, Dahlin AB et al.
QCM-D studies of human norovirus VLPs binding to glycosphingolipids in supported lipid bilayers reveal strain specific characteristics…
Kirjan sisällössä on
(1) taustaa NORO-viruksesta, senhistoriaa ja kuvataan kliinisiä ilmenemismuotoja, akuutin infektion merkit, kroonisen infektion merkit, epätavalliset kliiniset ilmenemät ja asymptomaattiset ( oireettomat) infektiot.
Sitten luokitellaan NORO-virukset ja kerrotaan niitten laboratoriodiagnostiikasta, epidemiologiasta, kausiluontoisesta ilmenemisestä ja tartuntatavoista. Sitten kerrotaan, miten virus lisääntyy soluviljelmissä, minkälainen on sen rakenne ja genomi, kuvataan viruskapsidia ja viruksen kaltaisia partikkeleita (VLPs). Sitten on vuorossa patogeneesin kuvaus. Selvitetään eläinmallit, joissa virusta voi tutkia (Murine norovirus, hiiren NORO-virus; ihmisen NORO-virus possuissa ja vasikoissa), Kuvataan NORO-virus zoonoosit.
Selvitetään NORO-viruksen luomaa immuniteettia ja vasta-aineitten ristireaktioita.
(2) Sitten keskitytään GLYKOBIOLOGIAAN, mikä on NORO-viruksen suhteen olennainen asia tietää.
Minkälainen on glykaanin biosynteesi? Kuvataan eräitä glykokonjugaatteja, glykoproteiineja, proteoglykaaneja ja glykosfingolipidejä.
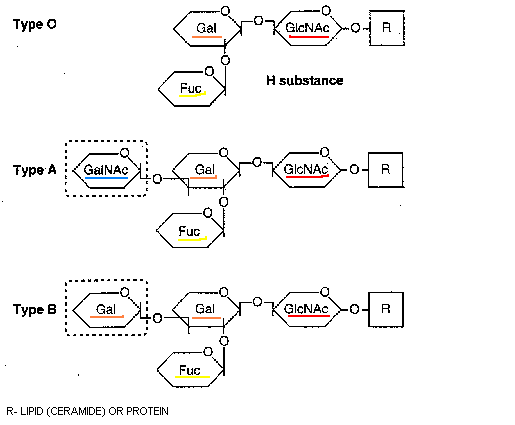
Nyt keskitytään erityisesti kudos-veriryhmäominaisuuksiin. Selvitetään ABO(H) ja Lewis histo-veriryhmäsysteemit. Kerrotaan näitten kudos-veriryhmille tyypillisten epitooppien biosynteesistä. Näitä ovat H-antigeenit, A-antigeenit, B-antigeenit ja Lewis antigeenit sekä sialyloidut ja sulfatoidut antigeenit. Sitten kirjoittaja selvittää, mikä on ABO(H) ja Lewis kudos-veriryhmä-järjestelmän varsinainen funktio, toiminta. Hän kuvaa ensiksi sialyyli Lewis x epitooppia ja sitten ABO(H) kudosveriryhmäjärjestelmää.
(3) Tutkija kertoo reseptoreista ja liittymistekijöistä.
(KOMMENTTI: Ihmisen solujen epitooppirakenteet toimivat virukselle reseptorina, joihin viruksilla on kehitetty ja jalostettu kykynsä takertua. Tutkija yrittää löytää sellaisia molekyylejä, jotka voisivatkin suojata ihmisen moninaisia epitooppeja niin, että virus ei saa niihin otetta)
Tutkija kuvaa glykokonjugaatteja,joita ovat proteoglykaanit .
Sitten selvitetään mitä on glykoproteiinit ja glykosfingolipidit.
Sitten kuvataan reseptoreita, eri viruslajeja ja virusten tropismia (= eli mikä virus valitsee minkäkinlaisen reseptorin itselleen kiinnittyessään ihmisen soluihin).
Mutta keholla on myös ”luonnollisia houkutuslintureseptoreitakin” ( natural decoy receptors) ja niiden tarkoitus on suojata soluja mikrobeilta fysikaalisena esteenä ja koska ne ovat lima- eli musiinirakenteisia ja geelimäisiä ja sakkaavat mikrobeja, ne poistuessaan siistivät soluympäristöä mikrobeista.
Proteiini-hiilihydraatti-interaktiot ovat multivalenttisia ja voivat muodostaa palautumattomia sidoksia.
Monet vaipattomat virukset luottavatkin multivalenttisuuteen liittyessään.
Glykaanit voivat moduloitua myös muullakin tavalla kuin multivalenttisuudella. Esim N-glykaanin terminaalisen epitoopin muuttuminen ja konformaatio vaikuttuu, jos lisätään ydintä, jossa on fukoosia ja haaroittuvaa GlcNAc. Glykaanin biologinen -tunnistuskyky vaikuttuu.
Mainitaan erilaisista yhteyksistä isäntäkehon geneettisten ominaisuuksien ja eri infektioiden kesken.
Näistä protektiivisuutta määrävistä piirteistä tunnetaan esim:
P. falciparum malaria ja Hemoglobiini sirppisoluanemiassa.
P. vivax malaria ja Duffy antigeenireseptori kemokiineille (DARC) ( Reseptorin puute) .
Malaria ja erytrosyytin kalvon juova 3.
HIV-1 ja kemokiinireseptori 5, CCR5. ( Reseptorin puute)
Creuzfeldt-Jacob prionitauti ja prioniproteiini PRPN.
NORO-virus ja FUT2 ( sekretorinen status) mainitaan tässä yhteydessä.
Nämä ovat kuusi vahvinta havaittua assosiaatiota infektoivan taudin ja yleisen geenipolymorfian kesken. Sekretorisen statuksen ja NORO-viruksen kesken on siis myös tällainen vahva assosiaatio havaittu.
Eräs tunnettu heikompi assosiaatio on niillä, joilla on P-veriryhmästä p fenotyyppi. Heillä on resistenssiä parvovirusta B19 kohtaan.
Eräs assosioaatio on O veriryhmän ja vaikean koleramuodon kesken.
Sitten tutkija kuvaa sekretorisen statuksen merkityksen infektioalttiudessa.
Esimerkkinä on monta patogeenia:
BAKTEERI ja sekretorinen status
E.coli tunnistaa glykosfingolipidia Gal alfa4Gal ym. eräitä globosarjan keramideja, joita esiintyi non-sekretorisilla yksilöillä ja ne ovat bakteeria vahvasti sitovia pintaepitooppeja.
Sekretorisilla yksilöillä biosynteesi menee kohti Globo H (tyyppi 4)-keramideja, jotka ovat huonoja bakteerin sitojia. Sekretorisilla on parempi suoja E-coli-virtsatulehduksia vastaan.
Non-sekretoriset yksilöt ovat alttiitta myös Haemofilus influenzae, Neisseria meningitidis ja Str. pneumoniae bakteereille.
VIRUS ja sekretorinen status
Päinvastoin kuin mainituissa bakteeritulehduksissa non-sekretoriset henkilöt ovat vähemmän edustettuja niitten joukossa, jotka kärsivät influenssa A tai B-viruksista, rhinoviruksesta, RSV ja ECHO viruksista. FUT2 genotyypillä ja HIV-1 progressiolla lienee myös yhteyttä, non-sekretorisilla taudin kulku on pitkällä ajalla non-progressiivinen.
Myös H-pylori infektiolla ja sekretorisella statuksella on yhteyttä (BabA adhesiini tunnistaa sekretorisesta geenistä riippuvan glykaanin). Sekretorinen status liittyy myös ei-infektiivisiin tauteihin. Non-sekretorisilla on plasman B12-pitoisuudet korkeammat kuin sekretorisilla.
(4) Tutkija selvittää, miten NORO-virus suhtautuu glykaaniin.
Tässä onkin erikoista historiaa. Kliiniset tutkimukset 1970- ja 1980 luvulta olivat hämmentäviä tarkattaessa Norwalk-virusvasta-aineitten ja taudille herkkyyden välistä yhteyttä. Erityisen hämmentävää oli se, että monilla yksilöillä, jotka olivat aina vastustuskykyisiä NORO-virusta vastaan, ei ilmennyt edes vasta-aineita sitä kohtaan.
Sitten kun vuonna 1982 havaittiin että NORO-virukselle vastustuskykyisyys oli suvuittain kulkeva seikka, alettiin arvella, että kyse on jostain geneettisestä asiasta. Kun pystyttiin tuottamaan menestyksekkäästi VLP hiukkasia NORO-viruksesta (1992), voitiin alkaa etsiä mihin reseptoreihin ne sitoututuvat. Vuonna 2000 saatiin oikein läpilyönti, kun RHDV:n osoitettiin sitoutuvan kudosveriryhmäantigeeneihin H tyyppiin 2 ja A typpiin 2. (RHDV on kaniinin verenvuototaudin virus. Tämä virus tunnisti alfa1,2 fukosyloituneita glykaaneja)
Kaksi vuotta edellisen löydön jälkeen sama tutkijaryhmä keksi, että myös Norwalk virus tunnistaa H-antigeenia, kun tehtiin Norwalk- virusaltistuksia ja VLP tutkimuksia . Norwalk virus tunnisti H-antigeenista tyypin 1 ja tyypin 3 ketjuja. Norwalk VLP:t tunnistivat sekretoripositiivisilta yksilöiltä gastroduodenaalisen (mahalukun ja pohjukaissuolen) liittymän epiteelisoluja, mutta ei non-sekretorisilta yksilöiltä. Tähän perustuu se vastustuskyky, mikä non-sekretorisilla henkilöillä on tätä virusta vastaan. (2003). Sitten pystyttiin ihmisen sekretorinen status genotyypittämään vuonna 2005. B-veriryhmäläiset olivat vastustuskykyisiä- nimittäin Norwalk VLP pystyi hemagglutinoimaan veriryhmän O, A ja AB punasoluja, mutta ei B-veriryhmän punasoluja.
Entä muut NORO-virukset?
NORO-virusten sitoutumista voi hyvin tutkia sylkinäytteistä, koska syljen musiinirakenteet muistuttavat gastrointestinaalisen kanavan epiteeleillä oleva musiiniglykaanirakennetta. On kerätty kaikista ABO(H) ja Lewis kudosveriryhmäläisistä sylkinäytteitä NORO-virustutkimuksiin. On saatu koottua tiedot sitoutumisista spesifisiin glykaaneihin, neoglykoproteiineihin tai polyakryyliamidiin. Koko joukko eri VLP kantoja tutkittiin. Geno-ryväs selvitettiin ja virukset testattiin non sekretorisilla ja sekretorisilla O, A ja B veriryhmä antigeenin omaavilla.
Vuonna 2004 havaittiin, että ABO(H) ja Lewis nimisiin kudosverirhmästruktuureihin kiinnittymisen lisäksi NORO-virus VLP löytää kiinnittymiskohtaa myös heparaanisulfaatista. Uusi löytö on, että GII VLP tunnistaa SLe x ja sen rakenneanalogeja.
(5) Taudinpurkauksista tehtyjä tutkimuksia
Ruotsissa vuonna 2005 selvitettiin eräs NORO-virusepidemia. 38 oireilevaa genotyypitettiin sekretorisen statuksen suhteen. Viruskanta oli GII.4, joka aiheuttaa sairaaloissa NORO-viruspurkauksia. Kolmesta sairaalan ulkopuolella tapahtuneesta infektionpurkaumasta diagnosoitiin GI.6 ja GII.6 tyypin NORO-viruskantoja. Yhteensä 53 symtomaattisesta ja 62 asymptomaattisesta yksilöstä katsottiin genotyyppi, FUT2 polymorfismi ja todettiin lopuksi, että oireilevien joukossa ei ollut yhtään non-sekretorista yksilöä. Hetero- ja homotsygoottisten sekretoristen yksilöitten kesken ei ollut eroa. Nonsekretoristen vastustuskyky GII.4 NORO-viruskantaa vastaan on todettu Tanskassakin (2007), mutta he voivat oireilla GI.3 kannasta. A-veriryhmän omaavia oli suurin joukko infektoituneista verrattuna asymptomaattisten määrään erään kiinalaisen tutkimuksen mukaan (2008). O-veriryhmäläisllä jakaantuma oli päinvastainen. GII.4 VLP tunnistaa sylkinäytteitä O-veriryhmäläisiltä huonommin kuin A, B ja AB-veriryhmäläisiltä.
Rintamaidon protektiivinen vaikutus
Rintamaito sisältää kudos-veriryhmäantigeenejä vapaina glykaaneina, glykosfingolipideinä (GSL) ja glykoproteiineina. Koeputkitutkimuksissa on havaittu, että sekretoristen (mutta ei nonsekretoristen) äitien maito estää NORO-viruksen VLP:itten sitoutumisen alfa1,2fukosyloituihin glykaaneihin. Fenotyyppisissä tutkimuksissa on havaittu, että veriryhmien A ja B antigeeneja ei ilmene maidossa. (Analogisesti tämän kanssa ei havaittu mitään blokeeraavaa vaikutusta eräällä sellaisella viruskannalla, joka tunnisti vain A ja B antigeenin sekretorisilla yksilöillä). Vauvojen ruokinta äidinmaidolla antaa suojaa kaliki-viruksia vastaan, esittaa eräs meksikolainen tutkimus. Kalikivirusinfektioita tapahtui vähemmän niillä vauvoilla, joiden saamassa maidossa oli korkeampi pitoisuus alfa1,2-linkkiytyneitä fukooseja.
Viruksen hiilihydraattia sitovat kohdat
Vasta-ainetutkimuksissa havaittiin, että vasta-ainetta sitova epitooppi lokalisoitui kapsidiproteiinin P2 domaaniin, mikä vastannee soluun sitoutumisesta. P partikkeli ja P dimeeri pitivät yllä sitoutumisen spesifisyyttä. Norwalk viruksessa näyttää olevan kaksi lähekkäin sijaitsevaa sitovaa kohtaa.
Sitova kohta on taskumainen ja havaittiin kiderakenteeltaan eräästä GII.4 kannasta, sen P-dimeeristä, -kun selvitettiin sen kompleksia B-veriryhmän trisakkaridin kanssa (2007). Sitovaa taskua ympäröi aminohapot; alfa1,2-linkkiytynyt fukoosi oli tiukasti sitoutuneena interaktioon seitsemän aminohapon kanssa, jotka kaikki ovat mitä konservoiduimpia GII NORO-viruskannoissa. Terminaalinen alfaGal tähde oli myös sitoutuneena, mutta beetaGal-toimi vähän kuin telineenä. Vuonna 2008 pystyttiin määrittämään ne aminohapot, jotka osallistuvat glykaanin tunnistukseen.
Esim. H tyyppi 1 rakenteen alfafukoositähe ja A rakenteen alfaGalNAc tähde tekivät hydrofobisen interaktion viruksen tryptofaaniin(Trp-375). Mutta B-ryhmän rakenteen alfaGal ei tehnyt interaktiota, koska siinä on OH-ryhmä N-asetamidoryhmän sijasta. Näin H-tyyppi1 pentasakkaridi ja A trisakkaridi sitoutuivat samaan kohtaa, mutta eri tavalla. Sitäpaitsi alfaFuc näytti olevan minimaalisesti mukana tässä A trisakkaridin sitoutumisessa.
http://www.cryst.bbk.ac.uk/pps97/assignments/projects/emilia/abo.gif
http://bp3.blogger.com/_DZH2cmCoois/Rdyfsu4z71I/AAAAAAAABAQ/LKDzwoxCg1M/s400/box+8-02.jpg
(6) Antiviraali terapia
LIITTYMISEN ESTÄJÄT YLEENSÄ
-viruksen kiinnittymisen estäjät, multivalentit
-anti-adheesiomolekyylit, glykaani-mimeetit, selektiiniantagonistit, anti-adhesiinit, sulfoidut -polysakkaridit,
-patogeenin agglutinaation estäminen
-monovalentit kiinnittymisen estäjät
-reseptoriia sitovan taskun blokeeraajat
-uusien lääkkeiden luomisen aluetta …
MUUT ANTIVIRUSLÄÄKKEET YLEENSÄ
-proteaasit,
- käänteisen transkription estäjät
- interferoniterapiat
- influenssalääkkeet, amantadiini, neuraminidaasin estäjät
-Suurin osa näistä noin 40 antiviraalista lääkkeestä kohdistuu viruksen entsyymeihin.
ROKOTTEET YLEENSÄ
Rokotteita on tehokkaasti kehitelty mm isorokkoa, poliota, tuhkarokkoa, parotiittia ja vihurirokkoa vastaan. Ainakin 15 viruspatogeenia on saanut vastaansa rokotteita. Influenssarokotuksia tulee ottaa vuosittain, koska virus on erittäin nopeasti antigeeniään muunteleva. Jotta rokote tehoaisi, sen tulee aiheuttaa neutraloivien vasta-aineitten muodostumista virusta kohtaan. Koska taas sellaiset vasta-aineet suuntautuvat viruksen reseptoria sitovaa kohtaa vastaan, täytyy olla tietoa viruksen ja reseptorin välisestä interaktiosta kehiteltäessä rokotetta.
ANTI-NOROVIRUSTERAPIA
Viime aikoina muutama harva tutkimus on esittänyt viruksen liittymistä estäviä lääkeaineita NORO-viruksen suhteen.
Yksi tutkimus esitti inhibiittoreita, jotka perustuvat alfa 1,2-fukosyloituneihin glykaani-hydrogeeli-konjugaatteihin (2006) . Eräs toinen tutkimus seuloin 5000 lääkkeenkaltaisen pienen molekyylin joukosta esiin 14 molekyyliä, jotka pystyivät 50%:seen inhibitiotehoon.
Väitöskirjan tekijä sanoo, että useista syistä ruotsalaiset tutkijat pitävät NORO-virusta sopivana kohteena luotaessa antiadhesiivista terapiaa.
Ensinnäkin on jo osoitettu mahdollisuudet poistaa erilaisten NORO-viruksien ja alfa 1,2-fukosyloituneiden glykaanien välinen interaktio. Viruksen ja glykaanin interaktion estämisestä on jo esimerkkinä sekretoristen äitien vauvojen saama suoja gastroenteriitiltä (2004)
Toiseksi on haastavaa kehittää rokotetta NORO-virusta vastaan, koska immuniteetti on kompleksista ja antigeenit ovat kovin vaihtelevia eri kantojen kesken. Vuosikymmennen aikana tapahtunut GII.4 kannan evoluutio viittaa siihen, että on tarve uudesta rokotteesta aivan influenssarokotteen tapaan, sillä uusia epidemisiä variantteja tulee tuon tuosta.
Kolmanneksi NORO-virusinfektio soveltuu hyvin profylaktisen terapian kohteeksi, koska se aiheuttaa laajoja purkauksia tiheissä väestöissä kuten sairaaloitten sisällä tai risteilylaivoilla. Tämä on tärkeää siksikin, koska anti-adhesiivisten aineitten terapeuttinen teho on parhainta kun sitä annetaan ennen patogeenialtistusta, mikä on analogista neuraminidaasi-inhibiittoreille influenssan suhteen.
Lopuksi NORO-virus-inhibiittorin anto saattaa olla yksinkertaista, koska sen on toimittava mahasuolikanavan alueella ”decoy ” mekanismilla, houkuttelemassa virusta. Täten lääkettä ei tarvitse optimoida kehon soluihin otettavaksi tai kuljetettavaksi kehokudoksiin, ja tämä alentaa sivuvaikutuksia muualla kehossa.
http://images.google.se/images?gbv=2&hl=sv&client=firefox-a&rls=org.mozilla%3Aen-US%3Aofficial&sa=3&q=NORO+virus&btnG=S%C3%B6k+bilder
Siis kommenttini: Summa summarum. Ollaan odottamassa uutta pahempaa NORO-virusta Pohjolaan. Mitään rokotetta tai täsmälääkettä ei ole olemassa. Käsienpesu, joka on käytössä pandemista influenssaa estävänä ja muu vastaava hygienia voi auttaa välttämään NORO-virustakin. ( Pesu vedellä ja saippualla ensisijaisesti)
{kind=link}
{kind=link}
Inga kommentarer:
Skicka en kommentar