Paul Spearman, Eric O.Freed ( Editors)
Hiv Interactions with Host Cell proteins .
Springer.
Current Topics in Microbiology and Immunology. 2009, siis ei aivan
tuoreinta tietoa. ISBN 978- 3- 642- 02174-9
Paul Spearmanin kirjassa
mainitaan sivulla 103 että HIV-1 virus elinsyklissään luottaa
strategiassaan myriadeihin interaktioihin isäntäsoluproteiinien kanssa.
Tämä tilanne asettaa valtavan haasteen tieteen tekniikalle.
Kappaleen otsikkona on
Imaging of HIV/Host Protein Interaction, kirjoittajina C M Danielson et T J Hope
- Traditionaaliset biokemialliset lähestymistavat
proteiini-proteiini-interaktioiden luotaamiseksi ovat kapasiteetiltaan
rajoitettuja tutkimaan intaktin solun tilavuudellisia ja dynaamisia
vuorovaikutuksia.
- Mutta fluoresoivin kuvausmenetelmin voidaan kuitenkin tutkia
viruksen ja isäntäsolun proteiinien välisiä paikallistumisia ja
dynaamisia interaktioita. Monta uutta fluoresointiin perustuvaa
kuvaustekniikkaa on kehitelty viime vuosikymmenen aikana ja ne ovat
olleet hyödyksi kuvattaessa HIV-1 virusproteiinien vuorovaikutusta
isäntäsoluun.
- Tekniikat mitä tässä kirjassa otetaan esiin:( engl)Colocalization versus interaction; Probing association by energy transfer; Improving imaging condition by increasing sensitivity and resolution
- ESIMERKKEJÄ asioista mihin näillä uusilla menetelmillä on
kohdistuttu ja mitkä tässä kappaleessa otetaan esiin. A. Viruksen
sisäänmeno ( entry), B. Viruksen ulostulo ( exit) C. Säätely ja lisäproteiinit ( regulatory and accessory proteins)
JOHDANTO
mainitsee seuraavia seikkoja: Retrovirus HIV koodaa itselleen vain 15 proteiinia, mutta on kehittänyt huippuunsa
isäntäsolun monien funktioiden anastamiskeinot. Niillä se hankkii itselleen sopivan miljöön elinsyklinsä täydellistämiseksi. Sillä on olemassa
myriadit interaktiot, jotka
suuntaavat uudelleen ja orjuuttavat isäntäsolun funktiota päämääränä
onnistunut infektoiminen- ja isäntäsolun uupumus ja tuhoutuminen.Kun
koetetaan selvitellä erilaisia molekulaarisia kohteita, joita voisi
terapeuttisesti manipuloida, on tiedettävä
viruksen komponenttien lisäksi myös
löydetyt proteiini-interaktiot ovat merkityksellisiä vain, jos ne tapahtuvat in vivo.
Jos sytoplasmassa ollut proteiini voisi rakenteellisesti tehdäkin
interaktion tumaproteiinin kanssa, tällainen teoreettinen interaktio eri
soluaitioista peräisin olevien proteiinien kesken on fysiologisesti
irrelevanttia.
Biokemialliset tutkimukset voivat kyllä luonnehtia
koeputkessa (
in vitro) joukoittain interaktioita, mutta uudet kuvaustekniikat visualisoivat interaktioita luonnollisina ja elävässä solussa.
ELÄVÄN SOLUN KUVANAMISTEKNIIKAT
ovat minimaalisen invasiivisia lähestymistapoja kuvaamassa tarkemmin
interaktioitten dynamiikka. Biokemiallisissa menetelmissä
löydetyt proteiini-interaktiot ovat merkityksellisiä vain, jos ne tapahtuvat in vivo ( elävässä kehossa) .
Jos sytoplasmassa ollut proteiini voisi rakenteellisesti tehdäkin
interaktion tumaproteiinin kanssa, tällainen teoreettinen interaktio
eri soluaitioista peräisin olevien proteiinien kesken on fysiologisesti irrelevanttia. Biokemialliset tutkimukset voivat kyllä luonnehtia
koeputkessa (
in vitro) joukoittain interaktioita, mutta uudet kuvaustekniikat visualisoivat interaktioita luonnollisina ja elävässä solussa.
Kymmenen viime vuotta on lisännyt runsaasti tietoa ja tekniikkaa. Kirjaansa on
Paul Spearman ottanut näistä tekniikoista kolme (yllämainittua) tarkemmin kuvattavaksi.
Sanoja mitä tekniikkaa kuvaavissa kappaleissa vilahtelee on mm
fluorescent microscopy, high resolution fluorescent imaging,
laser scanning confocal microscopy,
deconvolution microscopy,
computer- based image restoration, digital cameras,
spinning disc ,
fluorescence resonance energy transfer (FRET),
bimolecular fluorescent complementation (BiFC),
stimulated emission depletion (STED),
photo-activated localization microscopy (PALM),
total internal reflection fluorescence (TIRF) microscopy ,
traditional epifluorescence microscopy , evanescent field,
jne jne
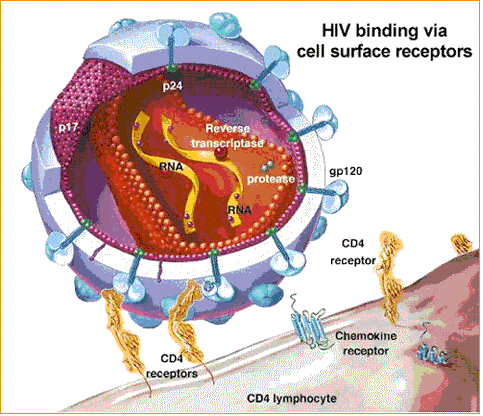
- A. Mitä löytöjä on tehty viruksen soluunmenovaiheesta?
HIV-1 viruksen
entry, sisäänmeno soluun. HIV menee CD4+Tyypin imusoluun sisälle sen jälkeen, kun virusvaipan (Envelope)
glykoproteiini gp120 on sitoutunut
CD4-reseptoriin, mikä sallii gp120 proteiinin tehdä interaktion ko-reseptoriin, joka on joko
CCR5 tai
CXCR4. Tällöin tapahtuu konformaation,
struktuurin muutos.
http://2009.igem.org/wiki/images/d/d1/HIV_absorb_cd4_and_ccr5.png
Tämä struktuurinmuutos tuo esiin hydrofobisen
fuusiopeptidin.
Fuusio viruksen
vaipan ja ihmissolun plasmakalvon kesken tapahtuu. Fuusiossa pääsee
viruksen sisällä oleva ydinkapsidi isäntäsolun sytoplasman
elinnesteitten alueelle.
http://img.medscape.com/fullsize/migrated/editorial/conferences/2001/325/retro.04.gif
Kapsidin seinämäosat purkautuvat ns vaipasta vapautumistapahtumassa
, uncoating -prosessissa ja sisällä oleva viruksen genominen aines, joka on kaksi
(+)plus sense RNA-säiettä alkaa muuttua
käänteiskopioivalla entsyymillä (RT) reversillä transkriptaasilla (RT), DNA- kielelle ja viruksesta hahmottuukin nyt
ds double strand DNA ja sitä kuljettava kompleksi PIC. (Sytoplasma ei siedä paljasta DNA:ta)
http://www.scielosp.org/img/revistas/aiss/v46n1/02f03.gif
http://www.nature.com/nrm/journal/v1/n1/images/nrm1000_040a_f1.gif
VirusRNA mallista valmistettu rusi tuore dsDNA pakkautuu ja kapaloituu PIC- hahmoiseksi
preintegraatiokompleksiksi,
joka vie vastasyntyneen DNA:n ja useita viruksen tuomia proteiineja
sekä solun proteiineja tumaan ja sitten tapahtuu alkuperäisin viraalin
koodin DNA-kielisen hahmon
integroituminen ihmisen kromosomistoon DNA:n joukkoon.
http://cochin.inserm.fr/research/scientific-departments/biocihp/team-berlioz-emiliani/nuclear-steps-of-hiv-1-replication/nuclear-steps-of-hiv-1-replication/images/Fig-1.jpg
Jos virus saa tehtyä tämän
DNA-
proviruksensa
, se antaa ikuiset merkit ihmisen genomiin. Tässä vaiheessa
infektiontekojärjestelmä voi sitten "hengähtää" ja olla latenttinakin,
piilevänä.
Entry-prosessi
luottaa hyvin lukuisiin interaktioihin isäntäsolun eri proteiinien
kanssa, mistä (häikäilemättömästä ryöstöpolitiikasta) virus hankkii
itselleen sekä ( tässä ja nyt-) hyödyn saada olla solun sytoplasmassa
tuhoutumatta sekä pääsyn ihmisen tumaan DNA:n alueelle ( varmistaen
lajitulevaisuuden) .
Näitten interaktioitten lokalisoitumisesta voidaan nykyisin helpommin saada tietoa
fluoresoivilla menetelmillä.
- Mobiilit reseptorit entry-vaiheessa
Normaalisti HIV-sisäänmenon reseptorit sijaitsevat tiheämpinä
tietyissä domaaneissa 10 nm toisistaan. Viruksen Envelope-vaippapinnan
gp120 indusoi interaktion reseptorien väliin.
FRET menetelmällä
2006 arvioitiin gp120 interaktio CD4 ja CCR5 reseptorien välillä.
Normaalisti nämä reseptorit muodostavat omat erilliset mikrodomaanit,
mutta jos on gp120 tulossa, ne tekevät kompleksin gp120:n kanssa
viruksen sisäänmenovaiheessa.
Gp120 saattaa CD4 ja CCR5 reseptorit yhteen elävän solun plasmamembraanissa Nämä reseptorit lokalisoituvat viruksen sisäänmenolle tärkeisiin
lipidilevykohtiin
ja jos nämä lipidilevyt kemiallisesti rikottiin, blokeerautui viruksen
gp120-indusoima signaali, mutta kolesterolin palauttaminen lipidilevyyn
ennallisti sen.
- Fluoresoivalla merkitsijällä merkattuja viruksia on tutkittu.
Ensimmäiset elävällä solulla tehdyt tutkimukset suoritettiin 2002, jolloin HIV viruksen lisäproteiini
Vpr oli merkattuna.Voitiin havaita, miten HIV menee soluihin ja
kaappaa solun mikrotubulaarisen verkoston käyttöönsä voidakseen suorittaa matkan kohti solun tumaa.
Virus oli tässä vaiheessa täysin riippuvainen solun
sytoskeletonrakenteesta saavuttaakseen virusgenomin lopullisen
päämäärän, ihmissolun tuman.
FRT menetelmällä on kuitenkin rajoituksensa. Esim.
Vpr
ilmeisesti katoaa kuvioista ennen kuin virusmateriaali on selviytynyt
tumaan asti, joten osasta HIV elämänsykliä ei voida kuvantaa tällä
merkitsijällä.
- Viruksen sisäänmenon (Entry) fuusiotapahtuman erottaminen endosytoosista
Eräs parannus virionin merkkaukseen on ollut Src:n 15
N-terminaalisen seriinin S15 merkkaus. Tämä virukseen inkorporoitunut
sígnaali katoaa fuusion jälkeen, mutta ei nonproduktiivisen reseptorista
riippumattoman endosytoosin jälkeen. S15 kohdistaa plasmamembraaniin ja
tämä sekvenssi inkorporoituu virioneihin. Kaksoismerkinnällä S15 ja
GFP-Vpr saadaan selkeä kuva virionista, joka on tullut soluun fuusiolla
ja on produktiivinen: viruksen sisäntulotapahtuma on produktiivinen.
Tällä erotetaan nonproduktiiviset, epärelevantit virionit.
Kaksoismerkintää voidaan käyttää
tutkittaessa retroviruksen restriktiotekijää rhTRIM5alfa, joka käy interaktioon sytoplasmisen HIV-kompleksin kanssa. TRIM5alfakompleksiin assosioituivat ne virionit, jotka olivat men
ettäneet S15-merkitsijäaineen ja tulleet isäntäsolun sytoplasmaan fuusiolla.
- Miten virus matkaa tumakeskukseen? IN- merkkaus , DNA
GFP-Vpr merkitsemisen rajoituksena on, että sillä voidaan kuvata
vain varhaisia postentry-tapahtumia, eikä ollenkaan tumassa
liikkumisia tai intrasellulaarisia HIV-kompleksin interaktioita.
Mutta yksi askel eteenpäin on ollut
integraasientsyymin (IN) merkkaus.
Vuonna 2006 se voitiin merkata (F1AsH-tagged integrase, fluorescein
arsenical hairpin) ja näin saatiin visualisoitua tumansisäisiä
liikkeitä.
Saatiin havaituksi HIV integraasin liikekinetiikasta
kohti tumaa sen käyttävän mikrotubuluksista ja aktiinista riippuvaa
liikkumista. HIV-kompleksit lokalisoituivat siinä systeemissä tumaan
rajoitetulla diffuusilla liikkeellä.
Vuonna 2008 F1AsH-menetelmää oli parannettu käyttämällä fluorophore-tagged integrase in trans Koska
paikalleenpistetyt sekvenssit provirussekvenssien sisällä usein
aiheuttavat prosessointiongelmia, tutkijat hyödynsivät sitä faktaa,
että Vpr on inkorporoituneena virioneihin ja
liittivät Vpr-tekijän fluoresoivasti merkattuun integraasiin.
Tämä tehtiin siten, että HIV-proteaasin ollessa läsnä pilkkoutuma
tapahtui niin, että Vpr erosi IN-EGFP:sta, merkatusta integraasista.
Täten Vpr-IN-EGFP ilmeni in trans proviruskonstruktion kanssa sisältäen
IN- deleetion sallien virionin produktion fluoresoivalla merkatulla
integraasilla.
Albanese,
jonka menetelmä tämä oli, tutki pikemminkin HIV viruksen suhdetta
isäntäsolun DNA:han eikä niinkään isäntäsolun proteiineihin. Kun
tutkittiin integroitumisen kohtia, paljastui
HIV viruksen taipumus ensisijaisesti integroitua aktiivisti transkriboituviin geeneihin.
Avoimen kysymyksenä on, mikä määrää tämän integroitumiskohdan
suosituimmuuden. He havaitsivat myös, että virionit eivät tapaa
vaellella pitkälle tumaan saapumisen jälkeen ja
suosituimmuudet on etsittävä purkautuneitten (dekondensoituneitten) kromatiinien alueelta.
Tämä on vasta löytöjen alkua siitä, mihin genomikohtiin HIV
assosioituu, mutta avaa monia näkymiä jatkotutkimuksiin tumansisäisistä
interaktioista.
- B. Entä miten virion lähtee solusta ulos (Exit)
Kun virus on integroinut sen käänteiskirjoitetun genominsa DNA:n isäntäsolun genomiin, muodostunut
provirus toimii templaattina, josta sitten voi tavalliseen tapaan transkriboitua RNA:ta
Nyt
alkaa sitten viruksen proteiinien syntetisoiminen ( HUOM: aivan kuin
solun omatkin proteiinit. Ensin lähettiRNA, mRNA, joka menee
sytoplasmaan ja etsii sopivat tRNA:t ja ribosomeilla translatoituu
peptidiketjuiksi ja proteiineiksi. Näitä proteiineja sitten kokoontuu
(Assembly) viruksen säätämässä sekvenssissä solun plasmakalvon lähelle
ja silmukoituu epäkypsinä virioneina infektoidusta solusta ulos. Tässä
on avuliaasti
apuna isäntäsolultta kaapatut järjestelmät:
endosomaalisen lajittelun kompleksi, jota virus tarvitsee tuotteen
kuljetuskoneistossa (ESCRT) kulkeutumiseen. Epäkypsät virionit
suorittavat kypsymisen ja sitten ovat valmiina infektoimaan muita
kohdesoluja.
Kuvantamistekniikat ovat antaneet paljon tärkeää uutta tietoa uusien virioneitten koostamisen interaktioista ja dynamiikasta.
Korreloiva kuvantamistekniikka ( Correlative Imaging). Fluoresoivalla merkkiaineella merkatut viriot sopivat
soluun menoa (entry) koskevien seikkojen tutkimuksen, mutta muita metodeita on käytettävä kuvaamaan nuppuilevan (budding) virionin
ulosmenoa (egres).Tässä
on kombinoitava tekniikkoja. Verrataan high-resolution electron
microscopy tekniikkaa ja fluoresoivaa kuvantamistekniikka
visualisoitaessa molekyylejä solun sisätilassa. Tällä voi visualisoida
retrovirusvirionin nuppuilun
Lisäksi voidaan hyödyntää transfektoidun Gag -polyproteiinin
multifotoni laser
scanning mikroskopiaa (MPM) ja jatkossa
scannin electron microscopy-tekniikkaa (
SEM). SEM voi nähdä yksittäiset nuppuilut, mutta on käytettävissä vain pinnan kuvaukseen.
Fluoresoiva tekniikka voi kuvantaa vain solun sisällä olevaa ja virionin silmukoitumisen voi vain päätellä
fluoresenssin katoamisesta.
On tutkittu myös yksittäisten partikkelien menemistä solun sisään
Larson et al. on
visualisoinut elävissä soluissa mm. yksittäisen HIV-1 viruksen
nuppuilun, virus like particles (VLP) ym. ja vertaillut fluoresoivissa
tekniikoissa saatuja tuloksia SEM tuloksiin.
- ESCRT eskortti mikrodomaanien kautta.Tetraspaniini
Aiemmin fluoresenssimikroskopialla oli havaittu HIV- partikkelin tekevän interaktion
tetraspaniinin
CD63 ja HIV Gag - polyproteiinin välille. Asian funktionaalista
merkitystä ei kuitenkaan voitu käsittää. Tavallisessa yksinkertaisessa
värjäyksessä vain pieni osa
CD63:sta
sijaitsee plasmakalvossa ja suurin osa sijaitsee intrasellulaarisissa
kalvoissa (2003) Seuraava askel tämän interaktion tutkimuksessa oli
vuodelta 2006. Interaktion jatkotutkimuksissa selektiivisesti värjätty
pinta-CD63 esiintyi ryväksittäin
mikrodomaaneissa eikä tasaisesti jakaantuneena kautta plasmakalvon.
Korkean resoluution immunoelektromikroskopialla kombinoituna fluoresoivaan kuvantamiseen voitiin nähdä elävässä solussa
tetraspaniinin rikastamat mikrodomaanit (TEM) .
TEM mikrodomaanit muodostuivat lähellä
clathriinilla katettuja alueita ja sytoskeletaalisia elementtejä.
Osoittautui, että
ESCRT1 komponentit
Tsg101 ja Vps28 (joita vaaditaan virionin nuppuiluun), rekrytoidaan
TEM:eille, joka sijaitsee samassa kuin Gag ( colocalization)
Vaikka on hyvin, hyvin paljon tutkittu HIV-1 virioneitten
ulospurkaantumista solusta, on asiassa varmaan jotakin löydettävää myös
fluoresenssi mikroskooppitutkimuksissa.
Traditionaalisissa co-lokalisaatiotutkimuksissa
havaittiin solukoneiston jaettua käyttöä viruksen ulospurkautumisen
ja solun sytokineesin kesken. Vuonna 2007 hiivassa havaittiin ESCRT -
koneiston ja sytokineesikoneiston komponenttien interaktioita j
a fluoresenssimikroskopialla tutkittiin näitten oletettujen interaktioiten paikallistumisia.
http://www.nature.com/nrmicro/journal/v5/n12/fig_tab/nrmicro1790_F2.html
ESCRT proteiinit Tsg101 ja Alix sitovat sentrosomaalista proteiinia Cep55,. joka osallistuu sytokineesiin. Kun Cep55 expressio katkistaan
siRNA:lla
eivät Tsg101 eikä Alix enää sijaitse tavallisilla paikoillaan.
Tutkijat kartoittivat Tsg101 aminohapot vastaamaan sitoutumisesta Cep55
proteiiniin ja havaitsivat deleetiolla tapahtuvan saman poikkeavan
lokalisoitumisen. Kun he havaitsivat tämän silmänpistävän esimerkin
viruksen tavasta anastaa niinkin tärkeä solun basaalifunktioitten kuin
solunjakautumisen koneisto , he löysivät Tsg101-Cep55 interaktion olevan välttämätön sytokineesille, mutta ei viruksen ulosnuppuilulle.
http://www.ncbi.nlm.nih.gov/pubmed/17853893
Tutkimalla
eri komponenttien vaatimuksia rekvisiittoina toimivissa solufunktioissa
jotka virus on valjastanut replikaatioonsa, saattaisi olla mahdollista
erotella spesifisiä interaktioita, jotka voitaisiin
antivirusterapialla keskeyttää säilyttämällä samalla tarvittavat
solutoiminnot.
- Virusten kaltaisten kappaleitten ulospurkaantumisen dynamiikkaa.
TIRF
(Total internal
reflection fluorescence ) on myös metodi virionmateriaalin kokoontumisen
ja ulospääsyn kuvantamisessa. Vuonna 2008 tehtiin TIRF kuvantamisen
kombinointi FRET:n kanssa ja f(uoresecnce recovery after
photobleaching) FRAP
- objektiivisempina metodeina interaktioitten määrittämiseksi viruksen purkaantumistutkimuksissa.
TIRF salli tutkijoitten kuvantaa
Gag VLP purkaantumista
reaaliajassa, mikä on vaikea tehtävä epifluoresenssissa, koska
sytoplasmisen Gag-peräisen signaalin vahvuus voittaa heikon
purkautumassa olevan VLP partikkelin signaalin. TIRF kuvantaa noin 100
nm alueelta.
Kinetiikkaa tutkittaessa
havaittiin kahta sorttia Gag-populaatiota. Oli hitaasti ilmaantuvaa ja
nopeasti ilmaantuvaa katoavaa. Käytettiin endosomaalisena merkitsijänä
CD63.
Nopeasti ilmenevä ja katoava Gag värjäytyi positiivisesti sekä CD63:lle
että clathriinille, joten tämä populaatio liittyi endosomeihin. Niinpä
he fokusoituivat hitaasti ilmenevään populaatioon, jota oli
plasmakalvossa eniten edustettuna. TIRF teki mahdolliseksi
karakterisoida näitä kineettisesti erilaisia Gag-populaatioita.
Sitten
he käyttivät FRET metodia tulkkaamaan hitaasti pintaan nousevia
partikkeleita Gag- molekyyleinä, jotka lähenevät toisiaan ja täten
ovat käymässä läpi kokoontumistaan viruksen kaltaisiksi partikkeleiksi
VLP.
Samantapaista FRET- metodiin perustuvaa menetelmää on
hyödynnetty Gag-polyproteiinin oligomerisaatiotutkimuksissa. sekä
elävässä solussa että viruksen kaltaisissa partikkeleissa (2004).
Vaikka
Juvenetin tutkimus (TIRF, 2008) kohdistui primääristi virionin
purkaantumisen dynamiikkaan, sitä voisi kokeilla lupaavana
lähestysmistapana luonnehdittaessa prosessin aikaisia interaktioita,
kuten soluproteiinien inkorporoitumista ja viraalin RNA:n
inkorporoitumista purkaantuvaan virioniin.
- Superresolution budding- nanometrien tarkkuutta
Betzig et al ( 2006) raportoivat
PALM menetelmän käytöstä (photoactivated localization microscopy). Tässä pystytään teoreettisesti erottamaan molekyylejä
muutaman nanometrin
tarkkuudella. Se ei lisää optista resoluutiota, mutta sallii
fluoresoivien signaalien etäisyyksien alajaksojen mittauksen.
Sekventiaalisella fotoaktivaatiolla ja subsekventiaalisella
photobleaching menetelmällä voidaan määritellä lukuisten molekyylien
sijaintia silloin kun muilla keinoilla ei päästä visualisoimaan
erikseen diffraktiorajan alla olevia tuhansia proteiineja (2006).
Signaalien keskuskohdat ja etäisyydet niiden välillä on
määritettävissä.
On myös
dual-color version tästä PALM teksniikasta.
Mikä merkitys tällaisella tarkuudella on?
Esim.
perinteisellä mikroskooppitekniikalla on havaittu "selvästi samaan
kohtaan lokalisoituvia proteiineja" ( colocalization), mutta nyt tämän
tekniikan ilmestyttyä voidaan osoittaa esim proteiinien olevan "hyvin
lähellä toisiaan, mutta eri molekyyli ryväksissä ( clusters) ".
Tällainen tekniikka saattaisi osoittautua arvokkaaksi, kun ollaan
ratkomassa viruksen ja isäntäsolun proteiinien välisiä oletettuja
interaktioita. Kun jatketaan tällä lisääntyvän tarkkuuden tiellä
nanometrisiä resoluutioita tavoitellen tullaan PALM-amalgamaatioon ja single-particle tracking metodiin, jota sanotaan
sptPALM (2008).
Hyötyä on
sptPALM menetelmän suuresta spatiaalisesta erotuskyvystä esim tutkittaessa
plasmamembraanin tuhansien yksittäisen molekyylien käytöstä
plasmamembraanin dynaaminen luonne visualisoituna. Kun käytetään
yksittäisten molekyylien selekiivistä fotoaktivaatiota voidaan
määritellä niitten vaikutusratoja hyvinkin pienillä solualueilla.
Manley et al.(2008) käyttivät
kombinoitua sptPALM
tekniikkaa Gag molekyylien jäljittämiseen plasmamembraanissa aivan
uskomattomalla tarkkuudella ja sitten he seurasivat eri
partikkelialaryhmien liikkeitä käyttäen
selektiivistä fotoaktivaatiota. Yksi kiinnostava löytö oli sekin, että Gag polyproteiinin liikkumaton osa pisti plasmamembraaniin.
Tämän oletettiin johtuvan interaktiosta tetraspaniinirunsaan
mikrodomaanin (TEM) kanssa. Täten Gag-dynamiikan eroavuuksia voitaneen
edelleen tutkia käyttämällä
dual colored PALM tekniikkaa tarkoituksella tutkia isäntäsoluproteiinien kanssa tapahtuvaa interaktiota.
- Mikä yhdistää Exit ja Entry tapahtumia?
Dendriittisoluilla (DC) on
kyky sitoa HIV tulematta produktiivisesti infektoituneeksi,
outo ilmiö, joka osaltaan vaikuttaa immuunivasteeseen ja samalla on
kuitenkin vielä yksi isäntäkehon funktio, jonka HIV on kaapannut. DC
solu voi sitoa ja hajoittaa HIV-virusta ja tuottaa vasta-aineita sitä
kohtaan, se voi myös sekvestroida virusta ja kuljettaa sitä T-soluille
ns trans-infektioprosessissa ja täten aiheuttaa osaltaan
systeemisen infektion puhkeamista.
Ratkaisematon kysymys on:
Missä tarkasti ottaen dendriittisolu varastoi virusta ennen kuin se toimittaa niitä pois T-solulle.
"Exosomi-malli"
olettaa että dendriittisolu varastoi virusta suojatuissa aitioissa
solun sisällä ja trans-infektoituminen pääsee tapahtumaan vain silloin
kun fuusiossa plasmasolun kanssa virion joutuu takaisin pintaan.
Uudet
tulokset osoittavat myös tätä vastustavan mallin olevan olemassa:
Oletetaan että internalisoituneet virionit ovat hajonneet lysosomeissa
ja vain pintaan pääseville virioneille on mahdollista aiheuttaa
trans-infektoituminen.
On näyttöä kummankin oletuksen
puolesta, mutta lopulta pystyttiin osoittamaan että virionit
varastoituvat internaaliseen aitioon, joka on taskumainen,
plasmamembraanin kanssayhteydessä oleva tila, josta käsin on
mahdollisuus päästä pintaan.Tämä siis vastustaa trans-infektiomallin
oletusta. Aktiinisytoskeleton, pinnasta rekrytoitu CD81 kertyivät samaan
aitioon kuin internaalinen HIV, jolle aktiinirakenne oli välttämätön
ulos pääsyyn.
Tällainen malli myös selittää, miksi solu pinnalle
applikoitu inhibiittori ja nestefaasinen merkitsijä pääsi virionin
aitioon. Samanlaista mekanismi on havaittu HIV leviämisessa
infektoituneesta solusta uuteen kohdesoluun. Konfiguraatiota sanotaan
virologiseksi synapsiksi. (2004)
Tästä on uudempaa tietoa:
http://jvi.asm.org/content/84/7/3516.short
infektoituneet
solut ovat paljon tehokkaampia HIV levittäjiä verrattaeessa soluttomiin
viruspartikkeliehin. Tätä pidettiin kauan mysteeriona, minkä nykyinen
kombinoitu kuvantamistekniikka vasta on ratkaissut. Sellaiset
interaktiot voivat olla kohtia, joissa plasmamembraani on suorassa
kontaktissa näiden kahden solun kesken: infektoituneen ja uuden
kohdesolun kesken. tai kontaktia välittää membraaniprojektiot kuten
sytoneemit ja nanotuubit.
Kaikissa tapauksissa infektiivisyys
stimuloituu, kun virus konsentroituu infektoituneesta viruksesta käsin
läheiseen kontaktiin kohdesolun fuusioon vaadittavien reseptoreitten
kanssa, jolloin uusi
entry, viruksen sisäänmeno, pääsee tapahtumaan suoraan
viruksen exit, ulosmenossa. .
- C. Säätely ja lisäproteiineista Tat ja Nef on myös uutta tietoa uudella tekniikalla.
Tat = transactivator of transcription protein of HIV
HIV viruksen transkriptiota trans-aktivoiva proteiini.
Se tehostaa viruksen RNA-synteesia vaikuttamalla trans-aktivaatiolle vastaavaan alueeseen
TAR (
trans-activation-responsive region) viruksen RNA:ssa. Se myös tekee interaktiota solukomponenteihin. Se tekee interaktionsa vaikuttamalla
P-TEBb tekijään, positiivisen transkription elongaasitekijään.
P-TEBb kinaasikomponentti sykliini T1
tekee kompleksin TAR kanssa HIV viruksen promoottorilla Tat ja
sykliiniT1 proteiinien interaktio tunnistettiin jo biokemiallisesti,
mutta vasta FRET-menetelmä pystyi ideaalisti kuvantamaan tätä oletettua
biokemiallista interaktiota ja määrittelemään sen fysiologisen
relevanssin: Tat vaikuttaa solun sisäiseen sykliini T1-lokalisaatioon.
Normaalist
i Sykliini T1 on tumatavara, kun taas
Tat tyypillisesti
asettuu diffuusisti kautta sekä nukleoplasman että tumahiukkasten
nucleolus. Kun niitä on samassa kohtaa, ne tekevät
uudelleenjärjestäytymisen. Tat vaikuttaa, että sykliini T1 asettuu toiseen paikkaan luonnollisesta akkumulaatiokohdastaan ja menee
transkriptiokohtaan ja samalla Tat ei enää olekaan diffuusisti, vaan a
kkumuloituu samaan
kohtaan. Sitten Tat ja sykliini T1 aktivoivat transkription, mikä oli
jo tiedettykin ennen uutta tekniikkaa. Ennen täydellistä
transkriptiota kuitenkin Tat irtautunee.
HIV viruksen säätelyproteiini
Nef
on vaadittava tekijä HIV infektion pahenemisessa AIDS asteelle. Nef
osallistuu säätämään vaimeaksi CD4 ja MCH-12 molekyylejä ihmisen
solupinnalta, mikä seikka lisää infektiivisyyttä. Tätä Nef vaikuttaa
tekemällä interaktioita monien ihmissoluproteiinien kanssa. Se tekee
interaktion
Hck proteiiniin ( proteiini-tyrosiinikinaasi), mikä seikka liittyy HIV patogeneesiin.
BiFC tekniikalla osoitettiin, että
Nef oligomerisaatio tapahtuu elävässä solussa, mikä taas on edellytys, jota kehon
Hck proteiini
voi aktivoitua . Odotetaan lisää tietoja Nef interaktiosta isäntäkehon
proteiinien kanssa. BiFC tekniikka on käytetty myös havaitsemaan
mahdollisia interaktioita retroviruksen Gag polyproteiinin ja aktiinin
kesken.
Tämän artikkelin kirjoittajat C M Danielsson ja T. Hope
toteavat, että on menossa oikein vallankumous HIV replikaatioon
osallistuvien solutekijöitten identifioimisessa.
siRNA ja shRNA
kirjastot poistogeenisistä solutekijöistä ovat johtaneet satojen
uusien tekijöitten multippeleitten ryhmien identifioimiseen ja niillä
voi olla interaktiota HIV proteiinien kanssa ja essentielliä osuutta HIV
biologiassa. Lähitulevaisuudessa tämä lista voi ainoastaan pidentyä.
Nykytekniikkaa
on pystynyt tuottamaan runsasta informaatiota näitten interaktioitten
funktionalisuudesta. Kuvantamisella saadaan valoa interaktioitten
paikallistumiseen ja dynamiikkaan ja komplisoitujenkin kysymysten
asettelujen ratkaisuja voidaan kombinoiduin metodein selvitellä.
Kehittämällä huippuunsa näitä jo korkealaatuisia tekniikoita (high
resolution imaging) (probing different stages of the viral life cycle)
aikamme tiedemiehet ovat hyvin varustautuneena vastaamaan kysymyksiin
HIV-1 virusproteiinien suhteesta ihmisen soluproteiinien miljööseen ja
täten luomaan perustaa sellaiselle tiedolle, jota voidaan terapian
luomiseen hyödyntää.
Päivitystä 23.2. 2015


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}