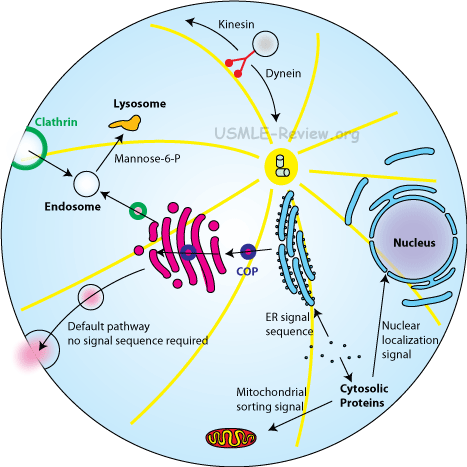
Adaptori proteiini (AP) kompleksit ovat avainaseman säätelijöitä sorteeraamassa sekretorisen ja endosomaalisen tien kuljetuksia.
AP kompleksit tunnistavat sorteeraussignaalit lastina olevan proteiinin (cargo proteins) sytoplasmisesta päädystä ja rekrytoivat soveliaita tukirakenneproteiineja tehokkaitten kuljetusrakkuloitten muodostamiseksi.
Ensiksi havaittiin kaksi AP kompleksia jotka saivat nimenkin AP-1 ja AP-2. Ne voitiin eristää clathriinin kattamista rakkuloista ja ne muodostivat clathrin- siltoja rakkuloihin. Niitä nimitettiin aluksi assembly proteins 1 and 2 nimillä (1987, 1983).
Sitten keksittiin vielä kaksi muuta AP-kompleksia AP-3 ja AP-4, kun tutkittiin imettäväisten cDNA kirjastoista AP-1 ja AP-2 sekvenssihomologioita. (1997, 1999).
Kaikkia neljää AP-kompleksiperhettä tavataan yleisesti sytosolisina heterotetrameereinä. Lisäksi AP-1 ja AP- 3 sisältävät solutyyppisiä spesifisiä isoformeja AP-1B esiintyy polarisoituneissa epiteelisoluissa ja AP-3B alayksikköineen esiintyy vain neuronaalisessa ja neuroendokriinisessä kudoksessa (1995).
Yksi isoista alayksiköistä asettuu interaktioon kalvon fosfatidyyli-inositolien (PI(4.5)P2 tai PI(3,4,5)P3 kanssa.
Toinen iso alayksikkö rekrytoi clathriinia käyttäen clathriinia sitovaa motiivia, jonka varassa se riippuu.
AP-1 ja AP-2 ovat runsaita clathriinilla katetuissa rakkuloissa, mutta AP-3 ja clathriinin välinen assosiaatio on epäselvä.
Biokemiallisesti on havaittu clathriinin puuttuvan AP-3:a sisältävistä rakkuloista. Mutta AP-3 alayksikkö beeta3 voi kuitenkin sitoa clathriinia.
Keskikokoiset alayksiköt tunnistavat sortteeraussignaaleja lastiproteiinin sytoplasmisesta päädystä ja tarjoavat myös kalvoankkurilisää sitoutumalla kalvon fosfatidyyli-inositoleihin.
http://usmle-review.org/cell-transport.gif
{kind=link}
Sortteeraussignaalit, jotka kiihdyttävät lajittelua clathriinilla-verhottuihin rakkuloihin, ovat tyrosiini(Y)- perusteisia signaaleja NPXY ja YXXPhi ja dileusiinipohjaisia signaaleja: DXXLL ja (DE)XXXL(LI). X on mikä tahansa aminohappo ja Phi on iso hydrofobinen ryhmä. Tiedetään että NPXY signaali on ainoa tunnettu, joka tekee interaktion monomeerisiin clathriiniadaptoreihin kuten autoresessiiviseen muotoon hyperkolesterolemiaproteiinia ARH.
Pienimmät alayksiköt omaavat rakenteellista tehtävää stabiloimalla heterotetrameerisen kompleksin ydintä. Mutta rakenteellisten ominaisuuksiensa lisäksi kaksi niistä osallistunee tunnistamaan dileusiiniperusteisia sorteeraussignaaleja,(DE)XXXL(LI), kuten HIV-1 viruksen nef-proteiinista ja LIMP-II proteiinista.
AP kompleksit osallistuvat proteiinien kuljettamiseen eri kohdissa post-Golgi verkostoa.
AP-1A välittää kahteen suuntaan (bidirectional) rakkulakuljetuksia trans-Golgi-verkoston(TGN) ja endosomien välillä.
AP-1B vastaa polarisoiduissa epiteelisoluissa rakkuloitten lajittelusta TGN:stä basolateraaliseen plasmakalvoon.
AP-2 lajittelee lastiproteiinia clathrin-välitteisiin endosyyttisiin rakkuloihin.
AP-3A on välttämätön lajittelija varhaisista endosomeista tai trans-Golgista (TGN) kohti myöhäisiä endosomeja (LE), MVB, lysosomeja ja lysosomin kaltaisia organelleja.
AP-3B arveltavasti toimii hermosoluissa synaptisten rakkuloitten muodostuksessa.
AP-4 osallistuu rakkuloitten sortteeraamiseen trans-Golgista lysosomeihin ja basolateraaliseen kuljetukseen transGolgista basolateraaliseen kalvoon.
AP kompleksit ovat osallistuneet myös HIV viruksen ja muitten vaipallisten virusten materiaalin kokoontumiseen (assembly) ja virioneitten nuppuiluun ulos isäntäsolusta( budding) .
On havaittu interaktio AP-3 ja HIV Gag polyproteiinin kesken. (2005)
AP-3 delta alayksikkö sitoutuu HIV-1 viruksen MA-alueen helix 1 kohtaan. Jos tämä AP-3-Gag interaktio katkeaa, muuntuu Gag polyproteiinin solun sisäinen asettautuminen ja samalla virionipartikkeleitten vapautuminen rajoittuu vahvasti (2005)
Gag-AP-3 interaktion katkeamsta Gag pystyy lokalisoitumaan vain minimaalisesti myöhäiseen endosomin LE/MVB aitioon) , mistä voi päätellä että juuri AP-3 johtaa Gag polyproteiinia tähän aitioon.
HIV-1 infektoituneissa dendriittisoluissa (DC solut virioneitten tuotanto vaati AP-3 kompleksia ja AP-3: :lla rikastuneita solunsisäisiä aitioita. AP-3:n rooli Gag polyproteiinin runsastuottoisessa kuljetuksessa ei ehkä yksistään rajoitu ainesten kokoontumiseen (assembly) intrasellulaarisen aition alueella. AP-3 voi kiihdyttää myös HIV-1 Gag polyproteiinin kuljetusta suoraan plasmakalvoon. On tarvetta yksimielisestä käsityksestä AP-3:n roolista HIV-assembly vaiheessa ja myös tarvitaan määrittelyä HIV-infekoituneessa makrofagissa tapahtuvasta AP-3 roolista.
Kun oli keksitty AP-3- Gag interaktio, havaittiin myös AP-2 ja HIV-1 Gag välillä linkkiä (2005). AP-2 kykeni sitoutumaan HIV-1 Gag polyproteiiniin tyrosiiniperusteiseen motiiviin MA-CA alueiten -liitoskohdassa. AP-2 - Gag interaktio kiihdytti partikkelin vapautumista.
Sitten keksittiin interaktio AP-1 ja HIV-1 Gag polyproteiinin MA- alueen välillä.
On mahdollista, että Gag polyproteiinin kuljetuksen suuntaamiseen osallistuu multippelit adaptoriproteiinikompleksit.
Myös HIV Env glykoproteiini (eräs muu HIV virionin komponenteista) on vahvasti osallistunut AP-kompleksi-interaktioihin.
Suurin vastaamaton kysymys on: MITEN Env saapuu assembly- kohtaan ja MITEN se assosioituu Gag polyproteiinin kanssa.
http://cochin.inserm.fr/research/scientific-departments/biocihp/team-berlioz-emiliani/late-steps-of-hiv-1-replication/late-steps-of-the-viral-replication/images/Fig-2-en.jpg
{kind=link}
Vaikka on alkanut olla selvää, että adaptoriproteiinikompleksit välittävät Env endosytoosia, ei ole kuitenkaan vahvistettu , miten tämä saattaisi osallistua tuomaan Gag ja Env yhteen (assembly). On pidetty mahdollisena, että Gag ja Env saavat co-transportin, yhteiskuljetuksen johonkin tavalliseen kokoontumispaikkaan; molemmat viruksen alayksiköt tekevät interaktion eri AP-alayksikön kanssa.
(Oma kommentti:
Tässä välillä pohdin , miksi solu lähettää HIV viruksen ulos aivan kuin se olisi solun oma erite. Johtuu tietysti siitä, että se tulee solun omasta DNA:sta, siis DNA provirus on pukeutunut solun DNA:n joukkoon, joten ei ehkä ole mahdollisuuksia tunnistaa mitään vierasta siinä ja se paketoidaan kuten kelpo tuote.
Sen jälkeen kun virus on tullut infektoituneen solun tuman lähettämänä mRNA muotona solukoneistoon sytoplasmaan, siinä ei ole enää tarpeeksi vieraan merkkiä, joten viruksen kätkemä oman genomin ohjelma koostua(assembly ) pääsee täysin toteutumaan ikään kuin se olisi solun oma assembly-funktio, koska viruksella ei ole sellaisia kykyjä. Se on ryöstänyt solun järjestelmän kåyttöönsä. Jos tässä vaiheessa yrittää kolaroida viruksen kanssa, siinä vahingoituu itse kehon ja ihmissolun järjestelmät. Strategian pitää olla aika snilleblixt, jotta tässä Gag polyproteiinin loppuvaiheessa tumasta mamebraaniin aikana kun se on polyproeiini- voi jotain vaikuttaa. Se vaikuttaa olevan turvassa kuin käenpoika pikkulinnun pesässä).
Nyt taas Spearmanin kirjan tekstiin:
TIP-47
On olemassa TIP-47. Tämä on adaptoriporteiinin kaltainen proteiini.
On havaittu että TIP-47 tekee interaktion sekä HIV Env että Gag proteiinien kanssa.
TIP-47 identifioitiin alunperin mannoosi-.6fosfatattireseptoreitetn (MPRs) sytoplasmisen domaanin sitoutumispartenerina ja essentiellinä välittäjänä myöhäisestä endosomista saapuvan MPR:n lajittelussa trans-Golgiverkostoon päin.
Vuonna 2003 havaittiin interaktio HIV Env ja TIP-47 kesken. Env pureutui TIP-47 molekyyliin YW -diaromaattisen motiivinsa avulla. Tämä motiivi on gp41 sytoplasmisessa päädyssä. Ja tästä interaktiosta saattoi seurata retrogradinen Env kuljetus plasmamembraanista trans-Golgiin(TGN).
Vuonna 2006 sama tiedemiesryhmä havaitsi että TIP-47 voi myös sitoutua Gag-polyproteiiniin sen MA-alueessa oleviin aminohappoihin 5- 16. , mikä osin kattaa samaa aluetta, mihin AP-3 pystyy tekemään interaktiota (H1 helix, aminohapot 11-19). Jos pyyhkäistään pois si-RNA vaimennuksella TIP-47- MA interaktio, virus oli huonompi infektiivisyydeltään ja Env-inkorporoitumisiltaan.
Tämä TIP-47- soluproteiinin mahdollinen rooli Gag-Env-adaptoriproteiinina on kiinnostava ja edelleen tutkittava asia, koska se saattanee päättäa viruksen välituotteiden koostamiskohdan.
HAKUSANA TIP-47, tail-interacting protein, 1 löytö
See 1 article found using an alternative search:
Source
Institut Cochin, Département Maladies Infectieuses, 27 Rue du Faubourg Saint Jacques, F-75014 Paris, France.
Abstract
The presence of the envelope glycoprotein Env in HIV-1 virions is essential for infectivity. To date, the molecular mechanism by which Env is packaged into virions has been largely unknown. Here, we show that TIP47 (tail-interacting protein of 47 kDa), which has been shown to interact with Env, also binds the MA (matrix) domain of HIV-1 Gag protein and that these three proteins form a ternary complex. Mutations in Gag that abrogate interaction with TIP47 inhibit Env incorporation and virion infectivity as well as colocalization between Gag and Env. We also show that TIP47 silencing impairs Env incorporation and infectivity and abolishes coimmunoprecipitation of Gag with Env. In contrast, overexpression of TIP47 increases Env packaging. Last, we demonstrate that TIP47 can interact simultaneously with Env and Gag. Taken together, our results show that TIP47 is a cellular cofactor that plays an essential role in Env incorporation, allowing the encounter and the physical association between HIV-1 Gag and Env proteins during the viral assembly process.
.
Inga kommentarer:
Skicka en kommentar