Peramivir
Systemaattinen (IUPAC) nimi
(1S,2S,3S,4R)-3-[(1S)-1-acetamido-2-ethyl-butyl]-4- (diaminomethylideneamino)-2-hydroxy-cyclopentane- 1-carboxylic acid
CAS number 229614-55-5
ATC code none, PubChem 151164
Kemiallinen kaava C15H28N4O4
Mol. mass 328.40722 g/mol
Bioavailability 100% (IV)
Metabolism ?
Puoliintumisaika 7.7 to 20.8 t jos munuaistoiminta on normaalia.
Erittyy munuaisten kautta.
Ei tiedetä miten lääke suhtautuu raskauteen. Metabolisia teitä ei tunneta tarpeeksi.
http://en.wikipedia.org/wiki/File:Peramivir.svg
PERAMIVIR on kokeellinen antiviruslääke, jonka on kehitellyt BioCryst Pharmaceuticals influenssan hoitoon. Se on laillistettu ensiapukäyttöön tietyillä sairaalapotilailla, joilla tiedetään olevan epäilty A H1N1 2009 influenssa.
PERAMIVIR on neuraminidaasi-inhibiittori, joka toimii transitiotilan analogisena neuraminidaasi-estäjänä influenssan neuraminidaasille ja sen takia pystyy estämään uusia viruksia purkautumasta infektoituneista solusita. PERAMIVIRIN kehittelyä ovat tukeneet
US Department of Health and Human Services osana USA:n hallituksen ponnisteluista inlfuenssapandemian uhkaa vastaan.
Tällä lääkkeellä on pitkä historia. Sen tablettimuoto hylättiin, koska biologinen saatavuus siitä oli huono (Johnson and Johnson). Mutta BioCryst Pharmaceuticals kehitteli injisoitavan muodon yhteistyössä E-Korean Green Cross Pharmaceuticals ja Japanin Shionogi Pharmaceuticals yhtiöitten kanssa. Lääke on nyt III vaiheen tutkimuksissa.
Lisänä netistä:
Resistenssitietoutta ennen pandemisen A h1n1 viruksen ilmestymistä
http://www.smittskyddsinstitutet.se/rav/metoder/influensa/diagnostik-och-resistensbestamning-betraffande-influensa-och-fagelinfluensa/
fredag 25 december 2009
{kind=link}
Zanamavir resistenssiä on havaittu A H1N1 viruksissa
Hurt AC, Holien JK, Parker M, Kelso A, Barr IG. Zanamivir-resistant influenza viruses with a novel neuraminidase mutation.J Virol. 2009 Oct;83(20):10366-73. Epub 2009 Jul 29. WHO Collaborating Centre for Reference and Research on Influenza, 10 Wreckyn St., North Melbourne, Victoria 3051, Australia.
Neuraminidaasin estäjäjiä ZANAMAVIR ja OSELTAMIVIR markkinoidaan influenssan hoitoon ja estoon ja niitä on kasoittan varastoituna moniin maihin pandemiassa käytettäväksi.
Vaikka tuoreissa katsauksissa oin havaittu melkoista lisääntymistä OSELTAMIVIR-resistentissä kausiluontoisessa A H1N1 influenssaviruksessa Euroopan, USA:n, Oceanian ja E-Afrikan alueilla, niin näihin asti ei ole merkittävästi raportoitu ZANAMAVIR-resistenssiä A H1N1 influenssaviruksissa tai muissakaan ihmisinfluenssaviruksissa.
Tässä artikkelissa tutkijat kertovat ottaneensa selvää OSELTAMIVIR- ja ZANAMAVIR-resistenssisstä kiertävissä kausiluontoisissa A H1N1- viruskannoissa, joita tavataan Australiassa ja kaakkois-Aasiassa. Kun analysoitiin 391 influenssa A H1n1 virusta 2006- 2008 väliseltä ajalta Australaasiasta ja Kaakkois-Aasiasta löydettiin yhdeksän virusta(2.3%) , jotka osoittivat huomattavsti alentunutta ZANAMAVIR-herkkyyttä ja niissä oli jo aiemmin kuvattu Gln136Lys (Q136K) neuraminidaasimutaatio.
Mutaatiolla ei ole mitään vaikutusta OSELTAMIVIR-herkkyyteen, muta se aiheutti noin 300-kertaisen ja 70 kertaisen alenemisen ZANAMAVIR- ja vastaavasti PERAMIVIR-herkkyydessä.
Q136K mutaation osuus ZANAMIVIR-resistenssissä varmistettiin käyttämällä käänteistä geneettistä tekniikkaa. Mielenkiintoista oli, että mutaatiota ei havaittu niissä primääreissä kliinisissä näytteissä, joista mutantti-isolaatit oli kasvatettu, mikä viittaisi siihen, että reistenttejä viruksia joko ilmenee hyvin pieni määrä primäärisissä näytteissä tai niitä muodostuu MDCK-solulinjojen viljelmien aikana.
Kun verrattiin herkkiin A H1N1 viruksiin niin Q136K-mutatoituneet kannat olivat parempikuntoisia viruksia kuin luonnolliset kannat MDCK soluissa, mutta niiden infektiivisyys ja tarttuvaisuus oli samanlaista ferretti koe-eläimissä.
Neuraminidaasin estäjäjiä ZANAMAVIR ja OSELTAMIVIR markkinoidaan influenssan hoitoon ja estoon ja niitä on kasoittan varastoituna moniin maihin pandemiassa käytettäväksi.
Vaikka tuoreissa katsauksissa oin havaittu melkoista lisääntymistä OSELTAMIVIR-resistentissä kausiluontoisessa A H1N1 influenssaviruksessa Euroopan, USA:n, Oceanian ja E-Afrikan alueilla, niin näihin asti ei ole merkittävästi raportoitu ZANAMAVIR-resistenssiä A H1N1 influenssaviruksissa tai muissakaan ihmisinfluenssaviruksissa.
Tässä artikkelissa tutkijat kertovat ottaneensa selvää OSELTAMIVIR- ja ZANAMAVIR-resistenssisstä kiertävissä kausiluontoisissa A H1N1- viruskannoissa, joita tavataan Australiassa ja kaakkois-Aasiassa. Kun analysoitiin 391 influenssa A H1n1 virusta 2006- 2008 väliseltä ajalta Australaasiasta ja Kaakkois-Aasiasta löydettiin yhdeksän virusta(2.3%) , jotka osoittivat huomattavsti alentunutta ZANAMAVIR-herkkyyttä ja niissä oli jo aiemmin kuvattu Gln136Lys (Q136K) neuraminidaasimutaatio.
Mutaatiolla ei ole mitään vaikutusta OSELTAMIVIR-herkkyyteen, muta se aiheutti noin 300-kertaisen ja 70 kertaisen alenemisen ZANAMAVIR- ja vastaavasti PERAMIVIR-herkkyydessä.
Q136K mutaation osuus ZANAMIVIR-resistenssissä varmistettiin käyttämällä käänteistä geneettistä tekniikkaa. Mielenkiintoista oli, että mutaatiota ei havaittu niissä primääreissä kliinisissä näytteissä, joista mutantti-isolaatit oli kasvatettu, mikä viittaisi siihen, että reistenttejä viruksia joko ilmenee hyvin pieni määrä primäärisissä näytteissä tai niitä muodostuu MDCK-solulinjojen viljelmien aikana.
Kun verrattiin herkkiin A H1N1 viruksiin niin Q136K-mutatoituneet kannat olivat parempikuntoisia viruksia kuin luonnolliset kannat MDCK soluissa, mutta niiden infektiivisyys ja tarttuvaisuus oli samanlaista ferretti koe-eläimissä.
söndag 20 december 2009
NORO-viruksen tieteellinen asema. Väitöskirja vuodelta 2009. Winter Vomiting Disease
Noroa norona NORO-viruksesta edelleen
NOROVIRUKSEN HOITOON SUUNNITELLAAN TERAPIAA.
NORO-viruksen tapoja ja sen vastaisen terapian hahmottelua käsittävä väitöskirja on antoisa ja koetan suomentaa siitä joitain kohtia, että saa käsitystä siitä, minkälaisen agenssin kanssa ollaan tekemisissä kun sanotaan että NORO-viruksesta on jälleen uutta uhkaa.
Väitöskirjan tekijä on Gustav E. Rydell et al. täällä Göteborgissa 2009.
Hänen väitöskirjansa nimi englanniksi on
Norovirus, causative agent of winter vomiting disease, exploits several histo-blood group glycans for adhesion.
Työ on tehty biomedisiinisessä instituutissa kliinisen kemian ja transfuusiolääketieteen osastolla Göteborgin Yliopistossa Sahlgrenskan Akatemiassa. Sahlgrenskan Keskuskirjastoon on kirja leimattu 5.11. 2009 sisään, joten asia on hyvin tuoretta.
Väitöskirjaa käsittää osatutkimukset, jotka tässä luettelen:
(I) Vuodelta 2006 Larsson MM, Rydell GEP et al.
Antibody prevalence and titer to norovirus (genotype II) correlate with secretor (FUT2) but not with ABO phenotype or Lewis (FUT3) genotype. The Journal of Infectious Diseases 194(10) pages 1422-7.Tästä olen aiemmin tehnyt lyhyttä mainintaa suomeksi.
(II) Vuodelta 2009: Rydell GE, Nilsson J et al.
Human noroviruses recognize sialyl Lewis x neoglycoprotein, Glycobiology 19(3), pages 309-20.
(III) Vuodelta 2009: Nilsson J, Rydell GE et al.
Norwalk-virus like particles bind specifically to A, H and difucosylated Lewis but not to B histo-blood group active glycosphingolipids. Glycoconjugate Journal, Apr 23 DOI 10.1007/s 10719-009-9237-x.
(IV) (2009) Rydell GE, Dahlin AB et al.
QCM-D studies of human norovirus VLPs binding to glycosphingolipids in supported lipid bilayers reveal strain specific characteristics…
Kirjan sisällössä on
(1) taustaa NORO-viruksesta, senhistoriaa ja kuvataan kliinisiä ilmenemismuotoja, akuutin infektion merkit, kroonisen infektion merkit, epätavalliset kliiniset ilmenemät ja asymptomaattiset ( oireettomat) infektiot.
Sitten luokitellaan NORO-virukset ja kerrotaan niitten laboratoriodiagnostiikasta, epidemiologiasta, kausiluontoisesta ilmenemisestä ja tartuntatavoista. Sitten kerrotaan, miten virus lisääntyy soluviljelmissä, minkälainen on sen rakenne ja genomi, kuvataan viruskapsidia ja viruksen kaltaisia partikkeleita (VLPs). Sitten on vuorossa patogeneesin kuvaus. Selvitetään eläinmallit, joissa virusta voi tutkia (Murine norovirus, hiiren NORO-virus; ihmisen NORO-virus possuissa ja vasikoissa), Kuvataan NORO-virus zoonoosit.
Selvitetään NORO-viruksen luomaa immuniteettia ja vasta-aineitten ristireaktioita.
(2) Sitten keskitytään GLYKOBIOLOGIAAN, mikä on NORO-viruksen suhteen olennainen asia tietää.
Minkälainen on glykaanin biosynteesi? Kuvataan eräitä glykokonjugaatteja, glykoproteiineja, proteoglykaaneja ja glykosfingolipidejä.
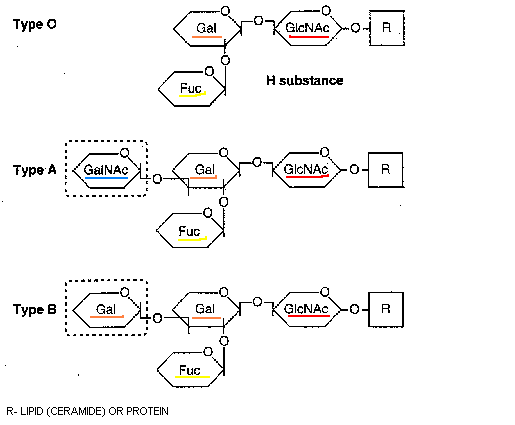
Nyt keskitytään erityisesti kudos-veriryhmäominaisuuksiin. Selvitetään ABO(H) ja Lewis histo-veriryhmäsysteemit. Kerrotaan näitten kudos-veriryhmille tyypillisten epitooppien biosynteesistä. Näitä ovat H-antigeenit, A-antigeenit, B-antigeenit ja Lewis antigeenit sekä sialyloidut ja sulfatoidut antigeenit. Sitten kirjoittaja selvittää, mikä on ABO(H) ja Lewis kudos-veriryhmä-järjestelmän varsinainen funktio, toiminta. Hän kuvaa ensiksi sialyyli Lewis x epitooppia ja sitten ABO(H) kudosveriryhmäjärjestelmää.
(3) Tutkija kertoo reseptoreista ja liittymistekijöistä.
(KOMMENTTI: Ihmisen solujen epitooppirakenteet toimivat virukselle reseptorina, joihin viruksilla on kehitetty ja jalostettu kykynsä takertua. Tutkija yrittää löytää sellaisia molekyylejä, jotka voisivatkin suojata ihmisen moninaisia epitooppeja niin, että virus ei saa niihin otetta)
Tutkija kuvaa glykokonjugaatteja,joita ovat proteoglykaanit .
Sitten selvitetään mitä on glykoproteiinit ja glykosfingolipidit.
Sitten kuvataan reseptoreita, eri viruslajeja ja virusten tropismia (= eli mikä virus valitsee minkäkinlaisen reseptorin itselleen kiinnittyessään ihmisen soluihin).
Mutta keholla on myös ”luonnollisia houkutuslintureseptoreitakin” ( natural decoy receptors) ja niiden tarkoitus on suojata soluja mikrobeilta fysikaalisena esteenä ja koska ne ovat lima- eli musiinirakenteisia ja geelimäisiä ja sakkaavat mikrobeja, ne poistuessaan siistivät soluympäristöä mikrobeista.
Proteiini-hiilihydraatti-interaktiot ovat multivalenttisia ja voivat muodostaa palautumattomia sidoksia.
Monet vaipattomat virukset luottavatkin multivalenttisuuteen liittyessään.
Glykaanit voivat moduloitua myös muullakin tavalla kuin multivalenttisuudella. Esim N-glykaanin terminaalisen epitoopin muuttuminen ja konformaatio vaikuttuu, jos lisätään ydintä, jossa on fukoosia ja haaroittuvaa GlcNAc. Glykaanin biologinen -tunnistuskyky vaikuttuu.
Mainitaan erilaisista yhteyksistä isäntäkehon geneettisten ominaisuuksien ja eri infektioiden kesken.
Näistä protektiivisuutta määrävistä piirteistä tunnetaan esim:
P. falciparum malaria ja Hemoglobiini sirppisoluanemiassa.
P. vivax malaria ja Duffy antigeenireseptori kemokiineille (DARC) ( Reseptorin puute) .
Malaria ja erytrosyytin kalvon juova 3.
HIV-1 ja kemokiinireseptori 5, CCR5. ( Reseptorin puute)
Creuzfeldt-Jacob prionitauti ja prioniproteiini PRPN.
NORO-virus ja FUT2 ( sekretorinen status) mainitaan tässä yhteydessä.
Nämä ovat kuusi vahvinta havaittua assosiaatiota infektoivan taudin ja yleisen geenipolymorfian kesken. Sekretorisen statuksen ja NORO-viruksen kesken on siis myös tällainen vahva assosiaatio havaittu.
Eräs tunnettu heikompi assosiaatio on niillä, joilla on P-veriryhmästä p fenotyyppi. Heillä on resistenssiä parvovirusta B19 kohtaan.
Eräs assosioaatio on O veriryhmän ja vaikean koleramuodon kesken.
Sitten tutkija kuvaa sekretorisen statuksen merkityksen infektioalttiudessa.
Esimerkkinä on monta patogeenia:
BAKTEERI ja sekretorinen status
E.coli tunnistaa glykosfingolipidia Gal alfa4Gal ym. eräitä globosarjan keramideja, joita esiintyi non-sekretorisilla yksilöillä ja ne ovat bakteeria vahvasti sitovia pintaepitooppeja.
Sekretorisilla yksilöillä biosynteesi menee kohti Globo H (tyyppi 4)-keramideja, jotka ovat huonoja bakteerin sitojia. Sekretorisilla on parempi suoja E-coli-virtsatulehduksia vastaan.
Non-sekretoriset yksilöt ovat alttiitta myös Haemofilus influenzae, Neisseria meningitidis ja Str. pneumoniae bakteereille.
VIRUS ja sekretorinen status
Päinvastoin kuin mainituissa bakteeritulehduksissa non-sekretoriset henkilöt ovat vähemmän edustettuja niitten joukossa, jotka kärsivät influenssa A tai B-viruksista, rhinoviruksesta, RSV ja ECHO viruksista. FUT2 genotyypillä ja HIV-1 progressiolla lienee myös yhteyttä, non-sekretorisilla taudin kulku on pitkällä ajalla non-progressiivinen.
Myös H-pylori infektiolla ja sekretorisella statuksella on yhteyttä (BabA adhesiini tunnistaa sekretorisesta geenistä riippuvan glykaanin). Sekretorinen status liittyy myös ei-infektiivisiin tauteihin. Non-sekretorisilla on plasman B12-pitoisuudet korkeammat kuin sekretorisilla.
(4) Tutkija selvittää, miten NORO-virus suhtautuu glykaaniin.
Tässä onkin erikoista historiaa. Kliiniset tutkimukset 1970- ja 1980 luvulta olivat hämmentäviä tarkattaessa Norwalk-virusvasta-aineitten ja taudille herkkyyden välistä yhteyttä. Erityisen hämmentävää oli se, että monilla yksilöillä, jotka olivat aina vastustuskykyisiä NORO-virusta vastaan, ei ilmennyt edes vasta-aineita sitä kohtaan.
Sitten kun vuonna 1982 havaittiin että NORO-virukselle vastustuskykyisyys oli suvuittain kulkeva seikka, alettiin arvella, että kyse on jostain geneettisestä asiasta. Kun pystyttiin tuottamaan menestyksekkäästi VLP hiukkasia NORO-viruksesta (1992), voitiin alkaa etsiä mihin reseptoreihin ne sitoututuvat. Vuonna 2000 saatiin oikein läpilyönti, kun RHDV:n osoitettiin sitoutuvan kudosveriryhmäantigeeneihin H tyyppiin 2 ja A typpiin 2. (RHDV on kaniinin verenvuototaudin virus. Tämä virus tunnisti alfa1,2 fukosyloituneita glykaaneja)
Kaksi vuotta edellisen löydön jälkeen sama tutkijaryhmä keksi, että myös Norwalk virus tunnistaa H-antigeenia, kun tehtiin Norwalk- virusaltistuksia ja VLP tutkimuksia . Norwalk virus tunnisti H-antigeenista tyypin 1 ja tyypin 3 ketjuja. Norwalk VLP:t tunnistivat sekretoripositiivisilta yksilöiltä gastroduodenaalisen (mahalukun ja pohjukaissuolen) liittymän epiteelisoluja, mutta ei non-sekretorisilta yksilöiltä. Tähän perustuu se vastustuskyky, mikä non-sekretorisilla henkilöillä on tätä virusta vastaan. (2003). Sitten pystyttiin ihmisen sekretorinen status genotyypittämään vuonna 2005. B-veriryhmäläiset olivat vastustuskykyisiä- nimittäin Norwalk VLP pystyi hemagglutinoimaan veriryhmän O, A ja AB punasoluja, mutta ei B-veriryhmän punasoluja.
Entä muut NORO-virukset?
NORO-virusten sitoutumista voi hyvin tutkia sylkinäytteistä, koska syljen musiinirakenteet muistuttavat gastrointestinaalisen kanavan epiteeleillä oleva musiiniglykaanirakennetta. On kerätty kaikista ABO(H) ja Lewis kudosveriryhmäläisistä sylkinäytteitä NORO-virustutkimuksiin. On saatu koottua tiedot sitoutumisista spesifisiin glykaaneihin, neoglykoproteiineihin tai polyakryyliamidiin. Koko joukko eri VLP kantoja tutkittiin. Geno-ryväs selvitettiin ja virukset testattiin non sekretorisilla ja sekretorisilla O, A ja B veriryhmä antigeenin omaavilla.
Vuonna 2004 havaittiin, että ABO(H) ja Lewis nimisiin kudosverirhmästruktuureihin kiinnittymisen lisäksi NORO-virus VLP löytää kiinnittymiskohtaa myös heparaanisulfaatista. Uusi löytö on, että GII VLP tunnistaa SLe x ja sen rakenneanalogeja.
(5) Taudinpurkauksista tehtyjä tutkimuksia
Ruotsissa vuonna 2005 selvitettiin eräs NORO-virusepidemia. 38 oireilevaa genotyypitettiin sekretorisen statuksen suhteen. Viruskanta oli GII.4, joka aiheuttaa sairaaloissa NORO-viruspurkauksia. Kolmesta sairaalan ulkopuolella tapahtuneesta infektionpurkaumasta diagnosoitiin GI.6 ja GII.6 tyypin NORO-viruskantoja. Yhteensä 53 symtomaattisesta ja 62 asymptomaattisesta yksilöstä katsottiin genotyyppi, FUT2 polymorfismi ja todettiin lopuksi, että oireilevien joukossa ei ollut yhtään non-sekretorista yksilöä. Hetero- ja homotsygoottisten sekretoristen yksilöitten kesken ei ollut eroa. Nonsekretoristen vastustuskyky GII.4 NORO-viruskantaa vastaan on todettu Tanskassakin (2007), mutta he voivat oireilla GI.3 kannasta. A-veriryhmän omaavia oli suurin joukko infektoituneista verrattuna asymptomaattisten määrään erään kiinalaisen tutkimuksen mukaan (2008). O-veriryhmäläisllä jakaantuma oli päinvastainen. GII.4 VLP tunnistaa sylkinäytteitä O-veriryhmäläisiltä huonommin kuin A, B ja AB-veriryhmäläisiltä.
Rintamaidon protektiivinen vaikutus
Rintamaito sisältää kudos-veriryhmäantigeenejä vapaina glykaaneina, glykosfingolipideinä (GSL) ja glykoproteiineina. Koeputkitutkimuksissa on havaittu, että sekretoristen (mutta ei nonsekretoristen) äitien maito estää NORO-viruksen VLP:itten sitoutumisen alfa1,2fukosyloituihin glykaaneihin. Fenotyyppisissä tutkimuksissa on havaittu, että veriryhmien A ja B antigeeneja ei ilmene maidossa. (Analogisesti tämän kanssa ei havaittu mitään blokeeraavaa vaikutusta eräällä sellaisella viruskannalla, joka tunnisti vain A ja B antigeenin sekretorisilla yksilöillä). Vauvojen ruokinta äidinmaidolla antaa suojaa kaliki-viruksia vastaan, esittaa eräs meksikolainen tutkimus. Kalikivirusinfektioita tapahtui vähemmän niillä vauvoilla, joiden saamassa maidossa oli korkeampi pitoisuus alfa1,2-linkkiytyneitä fukooseja.
Viruksen hiilihydraattia sitovat kohdat
Vasta-ainetutkimuksissa havaittiin, että vasta-ainetta sitova epitooppi lokalisoitui kapsidiproteiinin P2 domaaniin, mikä vastannee soluun sitoutumisesta. P partikkeli ja P dimeeri pitivät yllä sitoutumisen spesifisyyttä. Norwalk viruksessa näyttää olevan kaksi lähekkäin sijaitsevaa sitovaa kohtaa.
Sitova kohta on taskumainen ja havaittiin kiderakenteeltaan eräästä GII.4 kannasta, sen P-dimeeristä, -kun selvitettiin sen kompleksia B-veriryhmän trisakkaridin kanssa (2007). Sitovaa taskua ympäröi aminohapot; alfa1,2-linkkiytynyt fukoosi oli tiukasti sitoutuneena interaktioon seitsemän aminohapon kanssa, jotka kaikki ovat mitä konservoiduimpia GII NORO-viruskannoissa. Terminaalinen alfaGal tähde oli myös sitoutuneena, mutta beetaGal-toimi vähän kuin telineenä. Vuonna 2008 pystyttiin määrittämään ne aminohapot, jotka osallistuvat glykaanin tunnistukseen.
Esim. H tyyppi 1 rakenteen alfafukoositähe ja A rakenteen alfaGalNAc tähde tekivät hydrofobisen interaktion viruksen tryptofaaniin(Trp-375). Mutta B-ryhmän rakenteen alfaGal ei tehnyt interaktiota, koska siinä on OH-ryhmä N-asetamidoryhmän sijasta. Näin H-tyyppi1 pentasakkaridi ja A trisakkaridi sitoutuivat samaan kohtaa, mutta eri tavalla. Sitäpaitsi alfaFuc näytti olevan minimaalisesti mukana tässä A trisakkaridin sitoutumisessa.
http://www.cryst.bbk.ac.uk/pps97/assignments/projects/emilia/abo.gif
http://bp3.blogger.com/_DZH2cmCoois/Rdyfsu4z71I/AAAAAAAABAQ/LKDzwoxCg1M/s400/box+8-02.jpg
(6) Antiviraali terapia
LIITTYMISEN ESTÄJÄT YLEENSÄ
-viruksen kiinnittymisen estäjät, multivalentit
-anti-adheesiomolekyylit, glykaani-mimeetit, selektiiniantagonistit, anti-adhesiinit, sulfoidut -polysakkaridit,
-patogeenin agglutinaation estäminen
-monovalentit kiinnittymisen estäjät
-reseptoriia sitovan taskun blokeeraajat
-uusien lääkkeiden luomisen aluetta …
MUUT ANTIVIRUSLÄÄKKEET YLEENSÄ
-proteaasit,
- käänteisen transkription estäjät
- interferoniterapiat
- influenssalääkkeet, amantadiini, neuraminidaasin estäjät
-Suurin osa näistä noin 40 antiviraalista lääkkeestä kohdistuu viruksen entsyymeihin.
ROKOTTEET YLEENSÄ
Rokotteita on tehokkaasti kehitelty mm isorokkoa, poliota, tuhkarokkoa, parotiittia ja vihurirokkoa vastaan. Ainakin 15 viruspatogeenia on saanut vastaansa rokotteita. Influenssarokotuksia tulee ottaa vuosittain, koska virus on erittäin nopeasti antigeeniään muunteleva. Jotta rokote tehoaisi, sen tulee aiheuttaa neutraloivien vasta-aineitten muodostumista virusta kohtaan. Koska taas sellaiset vasta-aineet suuntautuvat viruksen reseptoria sitovaa kohtaa vastaan, täytyy olla tietoa viruksen ja reseptorin välisestä interaktiosta kehiteltäessä rokotetta.
ANTI-NOROVIRUSTERAPIA
Viime aikoina muutama harva tutkimus on esittänyt viruksen liittymistä estäviä lääkeaineita NORO-viruksen suhteen.
Yksi tutkimus esitti inhibiittoreita, jotka perustuvat alfa 1,2-fukosyloituneihin glykaani-hydrogeeli-konjugaatteihin (2006) . Eräs toinen tutkimus seuloin 5000 lääkkeenkaltaisen pienen molekyylin joukosta esiin 14 molekyyliä, jotka pystyivät 50%:seen inhibitiotehoon.
Väitöskirjan tekijä sanoo, että useista syistä ruotsalaiset tutkijat pitävät NORO-virusta sopivana kohteena luotaessa antiadhesiivista terapiaa.
Ensinnäkin on jo osoitettu mahdollisuudet poistaa erilaisten NORO-viruksien ja alfa 1,2-fukosyloituneiden glykaanien välinen interaktio. Viruksen ja glykaanin interaktion estämisestä on jo esimerkkinä sekretoristen äitien vauvojen saama suoja gastroenteriitiltä (2004)
Toiseksi on haastavaa kehittää rokotetta NORO-virusta vastaan, koska immuniteetti on kompleksista ja antigeenit ovat kovin vaihtelevia eri kantojen kesken. Vuosikymmennen aikana tapahtunut GII.4 kannan evoluutio viittaa siihen, että on tarve uudesta rokotteesta aivan influenssarokotteen tapaan, sillä uusia epidemisiä variantteja tulee tuon tuosta.
Kolmanneksi NORO-virusinfektio soveltuu hyvin profylaktisen terapian kohteeksi, koska se aiheuttaa laajoja purkauksia tiheissä väestöissä kuten sairaaloitten sisällä tai risteilylaivoilla. Tämä on tärkeää siksikin, koska anti-adhesiivisten aineitten terapeuttinen teho on parhainta kun sitä annetaan ennen patogeenialtistusta, mikä on analogista neuraminidaasi-inhibiittoreille influenssan suhteen.
Lopuksi NORO-virus-inhibiittorin anto saattaa olla yksinkertaista, koska sen on toimittava mahasuolikanavan alueella ”decoy ” mekanismilla, houkuttelemassa virusta. Täten lääkettä ei tarvitse optimoida kehon soluihin otettavaksi tai kuljetettavaksi kehokudoksiin, ja tämä alentaa sivuvaikutuksia muualla kehossa.
http://images.google.se/images?gbv=2&hl=sv&client=firefox-a&rls=org.mozilla%3Aen-US%3Aofficial&sa=3&q=NORO+virus&btnG=S%C3%B6k+bilder
Siis kommenttini: Summa summarum. Ollaan odottamassa uutta pahempaa NORO-virusta Pohjolaan. Mitään rokotetta tai täsmälääkettä ei ole olemassa. Käsienpesu, joka on käytössä pandemista influenssaa estävänä ja muu vastaava hygienia voi auttaa välttämään NORO-virustakin. ( Pesu vedellä ja saippualla ensisijaisesti)
NOROVIRUKSEN HOITOON SUUNNITELLAAN TERAPIAA.
NORO-viruksen tapoja ja sen vastaisen terapian hahmottelua käsittävä väitöskirja on antoisa ja koetan suomentaa siitä joitain kohtia, että saa käsitystä siitä, minkälaisen agenssin kanssa ollaan tekemisissä kun sanotaan että NORO-viruksesta on jälleen uutta uhkaa.
Väitöskirjan tekijä on Gustav E. Rydell et al. täällä Göteborgissa 2009.
Hänen väitöskirjansa nimi englanniksi on
Norovirus, causative agent of winter vomiting disease, exploits several histo-blood group glycans for adhesion.
Työ on tehty biomedisiinisessä instituutissa kliinisen kemian ja transfuusiolääketieteen osastolla Göteborgin Yliopistossa Sahlgrenskan Akatemiassa. Sahlgrenskan Keskuskirjastoon on kirja leimattu 5.11. 2009 sisään, joten asia on hyvin tuoretta.
Väitöskirjaa käsittää osatutkimukset, jotka tässä luettelen:
(I) Vuodelta 2006 Larsson MM, Rydell GEP et al.
Antibody prevalence and titer to norovirus (genotype II) correlate with secretor (FUT2) but not with ABO phenotype or Lewis (FUT3) genotype. The Journal of Infectious Diseases 194(10) pages 1422-7.Tästä olen aiemmin tehnyt lyhyttä mainintaa suomeksi.
(II) Vuodelta 2009: Rydell GE, Nilsson J et al.
Human noroviruses recognize sialyl Lewis x neoglycoprotein, Glycobiology 19(3), pages 309-20.
(III) Vuodelta 2009: Nilsson J, Rydell GE et al.
Norwalk-virus like particles bind specifically to A, H and difucosylated Lewis but not to B histo-blood group active glycosphingolipids. Glycoconjugate Journal, Apr 23 DOI 10.1007/s 10719-009-9237-x.
(IV) (2009) Rydell GE, Dahlin AB et al.
QCM-D studies of human norovirus VLPs binding to glycosphingolipids in supported lipid bilayers reveal strain specific characteristics…
Kirjan sisällössä on
(1) taustaa NORO-viruksesta, senhistoriaa ja kuvataan kliinisiä ilmenemismuotoja, akuutin infektion merkit, kroonisen infektion merkit, epätavalliset kliiniset ilmenemät ja asymptomaattiset ( oireettomat) infektiot.
Sitten luokitellaan NORO-virukset ja kerrotaan niitten laboratoriodiagnostiikasta, epidemiologiasta, kausiluontoisesta ilmenemisestä ja tartuntatavoista. Sitten kerrotaan, miten virus lisääntyy soluviljelmissä, minkälainen on sen rakenne ja genomi, kuvataan viruskapsidia ja viruksen kaltaisia partikkeleita (VLPs). Sitten on vuorossa patogeneesin kuvaus. Selvitetään eläinmallit, joissa virusta voi tutkia (Murine norovirus, hiiren NORO-virus; ihmisen NORO-virus possuissa ja vasikoissa), Kuvataan NORO-virus zoonoosit.
Selvitetään NORO-viruksen luomaa immuniteettia ja vasta-aineitten ristireaktioita.
(2) Sitten keskitytään GLYKOBIOLOGIAAN, mikä on NORO-viruksen suhteen olennainen asia tietää.
Minkälainen on glykaanin biosynteesi? Kuvataan eräitä glykokonjugaatteja, glykoproteiineja, proteoglykaaneja ja glykosfingolipidejä.
Nyt keskitytään erityisesti kudos-veriryhmäominaisuuksiin. Selvitetään ABO(H) ja Lewis histo-veriryhmäsysteemit. Kerrotaan näitten kudos-veriryhmille tyypillisten epitooppien biosynteesistä. Näitä ovat H-antigeenit, A-antigeenit, B-antigeenit ja Lewis antigeenit sekä sialyloidut ja sulfatoidut antigeenit. Sitten kirjoittaja selvittää, mikä on ABO(H) ja Lewis kudos-veriryhmä-järjestelmän varsinainen funktio, toiminta. Hän kuvaa ensiksi sialyyli Lewis x epitooppia ja sitten ABO(H) kudosveriryhmäjärjestelmää.
(3) Tutkija kertoo reseptoreista ja liittymistekijöistä.
(KOMMENTTI: Ihmisen solujen epitooppirakenteet toimivat virukselle reseptorina, joihin viruksilla on kehitetty ja jalostettu kykynsä takertua. Tutkija yrittää löytää sellaisia molekyylejä, jotka voisivatkin suojata ihmisen moninaisia epitooppeja niin, että virus ei saa niihin otetta)
Tutkija kuvaa glykokonjugaatteja,joita ovat proteoglykaanit .
Sitten selvitetään mitä on glykoproteiinit ja glykosfingolipidit.
Sitten kuvataan reseptoreita, eri viruslajeja ja virusten tropismia (= eli mikä virus valitsee minkäkinlaisen reseptorin itselleen kiinnittyessään ihmisen soluihin).
Mutta keholla on myös ”luonnollisia houkutuslintureseptoreitakin” ( natural decoy receptors) ja niiden tarkoitus on suojata soluja mikrobeilta fysikaalisena esteenä ja koska ne ovat lima- eli musiinirakenteisia ja geelimäisiä ja sakkaavat mikrobeja, ne poistuessaan siistivät soluympäristöä mikrobeista.
Proteiini-hiilihydraatti-interaktiot ovat multivalenttisia ja voivat muodostaa palautumattomia sidoksia.
Monet vaipattomat virukset luottavatkin multivalenttisuuteen liittyessään.
Glykaanit voivat moduloitua myös muullakin tavalla kuin multivalenttisuudella. Esim N-glykaanin terminaalisen epitoopin muuttuminen ja konformaatio vaikuttuu, jos lisätään ydintä, jossa on fukoosia ja haaroittuvaa GlcNAc. Glykaanin biologinen -tunnistuskyky vaikuttuu.
Mainitaan erilaisista yhteyksistä isäntäkehon geneettisten ominaisuuksien ja eri infektioiden kesken.
Näistä protektiivisuutta määrävistä piirteistä tunnetaan esim:
P. falciparum malaria ja Hemoglobiini sirppisoluanemiassa.
P. vivax malaria ja Duffy antigeenireseptori kemokiineille (DARC) ( Reseptorin puute) .
Malaria ja erytrosyytin kalvon juova 3.
HIV-1 ja kemokiinireseptori 5, CCR5. ( Reseptorin puute)
Creuzfeldt-Jacob prionitauti ja prioniproteiini PRPN.
NORO-virus ja FUT2 ( sekretorinen status) mainitaan tässä yhteydessä.
Nämä ovat kuusi vahvinta havaittua assosiaatiota infektoivan taudin ja yleisen geenipolymorfian kesken. Sekretorisen statuksen ja NORO-viruksen kesken on siis myös tällainen vahva assosiaatio havaittu.
Eräs tunnettu heikompi assosiaatio on niillä, joilla on P-veriryhmästä p fenotyyppi. Heillä on resistenssiä parvovirusta B19 kohtaan.
Eräs assosioaatio on O veriryhmän ja vaikean koleramuodon kesken.
Sitten tutkija kuvaa sekretorisen statuksen merkityksen infektioalttiudessa.
Esimerkkinä on monta patogeenia:
BAKTEERI ja sekretorinen status
E.coli tunnistaa glykosfingolipidia Gal alfa4Gal ym. eräitä globosarjan keramideja, joita esiintyi non-sekretorisilla yksilöillä ja ne ovat bakteeria vahvasti sitovia pintaepitooppeja.
Sekretorisilla yksilöillä biosynteesi menee kohti Globo H (tyyppi 4)-keramideja, jotka ovat huonoja bakteerin sitojia. Sekretorisilla on parempi suoja E-coli-virtsatulehduksia vastaan.
Non-sekretoriset yksilöt ovat alttiitta myös Haemofilus influenzae, Neisseria meningitidis ja Str. pneumoniae bakteereille.
VIRUS ja sekretorinen status
Päinvastoin kuin mainituissa bakteeritulehduksissa non-sekretoriset henkilöt ovat vähemmän edustettuja niitten joukossa, jotka kärsivät influenssa A tai B-viruksista, rhinoviruksesta, RSV ja ECHO viruksista. FUT2 genotyypillä ja HIV-1 progressiolla lienee myös yhteyttä, non-sekretorisilla taudin kulku on pitkällä ajalla non-progressiivinen.
Myös H-pylori infektiolla ja sekretorisella statuksella on yhteyttä (BabA adhesiini tunnistaa sekretorisesta geenistä riippuvan glykaanin). Sekretorinen status liittyy myös ei-infektiivisiin tauteihin. Non-sekretorisilla on plasman B12-pitoisuudet korkeammat kuin sekretorisilla.
(4) Tutkija selvittää, miten NORO-virus suhtautuu glykaaniin.
Tässä onkin erikoista historiaa. Kliiniset tutkimukset 1970- ja 1980 luvulta olivat hämmentäviä tarkattaessa Norwalk-virusvasta-aineitten ja taudille herkkyyden välistä yhteyttä. Erityisen hämmentävää oli se, että monilla yksilöillä, jotka olivat aina vastustuskykyisiä NORO-virusta vastaan, ei ilmennyt edes vasta-aineita sitä kohtaan.
Sitten kun vuonna 1982 havaittiin että NORO-virukselle vastustuskykyisyys oli suvuittain kulkeva seikka, alettiin arvella, että kyse on jostain geneettisestä asiasta. Kun pystyttiin tuottamaan menestyksekkäästi VLP hiukkasia NORO-viruksesta (1992), voitiin alkaa etsiä mihin reseptoreihin ne sitoututuvat. Vuonna 2000 saatiin oikein läpilyönti, kun RHDV:n osoitettiin sitoutuvan kudosveriryhmäantigeeneihin H tyyppiin 2 ja A typpiin 2. (RHDV on kaniinin verenvuototaudin virus. Tämä virus tunnisti alfa1,2 fukosyloituneita glykaaneja)
Kaksi vuotta edellisen löydön jälkeen sama tutkijaryhmä keksi, että myös Norwalk virus tunnistaa H-antigeenia, kun tehtiin Norwalk- virusaltistuksia ja VLP tutkimuksia . Norwalk virus tunnisti H-antigeenista tyypin 1 ja tyypin 3 ketjuja. Norwalk VLP:t tunnistivat sekretoripositiivisilta yksilöiltä gastroduodenaalisen (mahalukun ja pohjukaissuolen) liittymän epiteelisoluja, mutta ei non-sekretorisilta yksilöiltä. Tähän perustuu se vastustuskyky, mikä non-sekretorisilla henkilöillä on tätä virusta vastaan. (2003). Sitten pystyttiin ihmisen sekretorinen status genotyypittämään vuonna 2005. B-veriryhmäläiset olivat vastustuskykyisiä- nimittäin Norwalk VLP pystyi hemagglutinoimaan veriryhmän O, A ja AB punasoluja, mutta ei B-veriryhmän punasoluja.
Entä muut NORO-virukset?
NORO-virusten sitoutumista voi hyvin tutkia sylkinäytteistä, koska syljen musiinirakenteet muistuttavat gastrointestinaalisen kanavan epiteeleillä oleva musiiniglykaanirakennetta. On kerätty kaikista ABO(H) ja Lewis kudosveriryhmäläisistä sylkinäytteitä NORO-virustutkimuksiin. On saatu koottua tiedot sitoutumisista spesifisiin glykaaneihin, neoglykoproteiineihin tai polyakryyliamidiin. Koko joukko eri VLP kantoja tutkittiin. Geno-ryväs selvitettiin ja virukset testattiin non sekretorisilla ja sekretorisilla O, A ja B veriryhmä antigeenin omaavilla.
Vuonna 2004 havaittiin, että ABO(H) ja Lewis nimisiin kudosverirhmästruktuureihin kiinnittymisen lisäksi NORO-virus VLP löytää kiinnittymiskohtaa myös heparaanisulfaatista. Uusi löytö on, että GII VLP tunnistaa SLe x ja sen rakenneanalogeja.
(5) Taudinpurkauksista tehtyjä tutkimuksia
Ruotsissa vuonna 2005 selvitettiin eräs NORO-virusepidemia. 38 oireilevaa genotyypitettiin sekretorisen statuksen suhteen. Viruskanta oli GII.4, joka aiheuttaa sairaaloissa NORO-viruspurkauksia. Kolmesta sairaalan ulkopuolella tapahtuneesta infektionpurkaumasta diagnosoitiin GI.6 ja GII.6 tyypin NORO-viruskantoja. Yhteensä 53 symtomaattisesta ja 62 asymptomaattisesta yksilöstä katsottiin genotyyppi, FUT2 polymorfismi ja todettiin lopuksi, että oireilevien joukossa ei ollut yhtään non-sekretorista yksilöä. Hetero- ja homotsygoottisten sekretoristen yksilöitten kesken ei ollut eroa. Nonsekretoristen vastustuskyky GII.4 NORO-viruskantaa vastaan on todettu Tanskassakin (2007), mutta he voivat oireilla GI.3 kannasta. A-veriryhmän omaavia oli suurin joukko infektoituneista verrattuna asymptomaattisten määrään erään kiinalaisen tutkimuksen mukaan (2008). O-veriryhmäläisllä jakaantuma oli päinvastainen. GII.4 VLP tunnistaa sylkinäytteitä O-veriryhmäläisiltä huonommin kuin A, B ja AB-veriryhmäläisiltä.
Rintamaidon protektiivinen vaikutus
Rintamaito sisältää kudos-veriryhmäantigeenejä vapaina glykaaneina, glykosfingolipideinä (GSL) ja glykoproteiineina. Koeputkitutkimuksissa on havaittu, että sekretoristen (mutta ei nonsekretoristen) äitien maito estää NORO-viruksen VLP:itten sitoutumisen alfa1,2fukosyloituihin glykaaneihin. Fenotyyppisissä tutkimuksissa on havaittu, että veriryhmien A ja B antigeeneja ei ilmene maidossa. (Analogisesti tämän kanssa ei havaittu mitään blokeeraavaa vaikutusta eräällä sellaisella viruskannalla, joka tunnisti vain A ja B antigeenin sekretorisilla yksilöillä). Vauvojen ruokinta äidinmaidolla antaa suojaa kaliki-viruksia vastaan, esittaa eräs meksikolainen tutkimus. Kalikivirusinfektioita tapahtui vähemmän niillä vauvoilla, joiden saamassa maidossa oli korkeampi pitoisuus alfa1,2-linkkiytyneitä fukooseja.
Viruksen hiilihydraattia sitovat kohdat
Vasta-ainetutkimuksissa havaittiin, että vasta-ainetta sitova epitooppi lokalisoitui kapsidiproteiinin P2 domaaniin, mikä vastannee soluun sitoutumisesta. P partikkeli ja P dimeeri pitivät yllä sitoutumisen spesifisyyttä. Norwalk viruksessa näyttää olevan kaksi lähekkäin sijaitsevaa sitovaa kohtaa.
Sitova kohta on taskumainen ja havaittiin kiderakenteeltaan eräästä GII.4 kannasta, sen P-dimeeristä, -kun selvitettiin sen kompleksia B-veriryhmän trisakkaridin kanssa (2007). Sitovaa taskua ympäröi aminohapot; alfa1,2-linkkiytynyt fukoosi oli tiukasti sitoutuneena interaktioon seitsemän aminohapon kanssa, jotka kaikki ovat mitä konservoiduimpia GII NORO-viruskannoissa. Terminaalinen alfaGal tähde oli myös sitoutuneena, mutta beetaGal-toimi vähän kuin telineenä. Vuonna 2008 pystyttiin määrittämään ne aminohapot, jotka osallistuvat glykaanin tunnistukseen.
Esim. H tyyppi 1 rakenteen alfafukoositähe ja A rakenteen alfaGalNAc tähde tekivät hydrofobisen interaktion viruksen tryptofaaniin(Trp-375). Mutta B-ryhmän rakenteen alfaGal ei tehnyt interaktiota, koska siinä on OH-ryhmä N-asetamidoryhmän sijasta. Näin H-tyyppi1 pentasakkaridi ja A trisakkaridi sitoutuivat samaan kohtaa, mutta eri tavalla. Sitäpaitsi alfaFuc näytti olevan minimaalisesti mukana tässä A trisakkaridin sitoutumisessa.
http://www.cryst.bbk.ac.uk/pps97/assignments/projects/emilia/abo.gif
{kind=link}
http://bp3.blogger.com/_DZH2cmCoois/Rdyfsu4z71I/AAAAAAAABAQ/LKDzwoxCg1M/s400/box+8-02.jpg
{kind=link}
(6) Antiviraali terapia
LIITTYMISEN ESTÄJÄT YLEENSÄ
-viruksen kiinnittymisen estäjät, multivalentit
-anti-adheesiomolekyylit, glykaani-mimeetit, selektiiniantagonistit, anti-adhesiinit, sulfoidut -polysakkaridit,
-patogeenin agglutinaation estäminen
-monovalentit kiinnittymisen estäjät
-reseptoriia sitovan taskun blokeeraajat
-uusien lääkkeiden luomisen aluetta …
MUUT ANTIVIRUSLÄÄKKEET YLEENSÄ
-proteaasit,
- käänteisen transkription estäjät
- interferoniterapiat
- influenssalääkkeet, amantadiini, neuraminidaasin estäjät
-Suurin osa näistä noin 40 antiviraalista lääkkeestä kohdistuu viruksen entsyymeihin.
ROKOTTEET YLEENSÄ
Rokotteita on tehokkaasti kehitelty mm isorokkoa, poliota, tuhkarokkoa, parotiittia ja vihurirokkoa vastaan. Ainakin 15 viruspatogeenia on saanut vastaansa rokotteita. Influenssarokotuksia tulee ottaa vuosittain, koska virus on erittäin nopeasti antigeeniään muunteleva. Jotta rokote tehoaisi, sen tulee aiheuttaa neutraloivien vasta-aineitten muodostumista virusta kohtaan. Koska taas sellaiset vasta-aineet suuntautuvat viruksen reseptoria sitovaa kohtaa vastaan, täytyy olla tietoa viruksen ja reseptorin välisestä interaktiosta kehiteltäessä rokotetta.
ANTI-NOROVIRUSTERAPIA
Viime aikoina muutama harva tutkimus on esittänyt viruksen liittymistä estäviä lääkeaineita NORO-viruksen suhteen.
Yksi tutkimus esitti inhibiittoreita, jotka perustuvat alfa 1,2-fukosyloituneihin glykaani-hydrogeeli-konjugaatteihin (2006) . Eräs toinen tutkimus seuloin 5000 lääkkeenkaltaisen pienen molekyylin joukosta esiin 14 molekyyliä, jotka pystyivät 50%:seen inhibitiotehoon.
Väitöskirjan tekijä sanoo, että useista syistä ruotsalaiset tutkijat pitävät NORO-virusta sopivana kohteena luotaessa antiadhesiivista terapiaa.
Ensinnäkin on jo osoitettu mahdollisuudet poistaa erilaisten NORO-viruksien ja alfa 1,2-fukosyloituneiden glykaanien välinen interaktio. Viruksen ja glykaanin interaktion estämisestä on jo esimerkkinä sekretoristen äitien vauvojen saama suoja gastroenteriitiltä (2004)
Toiseksi on haastavaa kehittää rokotetta NORO-virusta vastaan, koska immuniteetti on kompleksista ja antigeenit ovat kovin vaihtelevia eri kantojen kesken. Vuosikymmennen aikana tapahtunut GII.4 kannan evoluutio viittaa siihen, että on tarve uudesta rokotteesta aivan influenssarokotteen tapaan, sillä uusia epidemisiä variantteja tulee tuon tuosta.
Kolmanneksi NORO-virusinfektio soveltuu hyvin profylaktisen terapian kohteeksi, koska se aiheuttaa laajoja purkauksia tiheissä väestöissä kuten sairaaloitten sisällä tai risteilylaivoilla. Tämä on tärkeää siksikin, koska anti-adhesiivisten aineitten terapeuttinen teho on parhainta kun sitä annetaan ennen patogeenialtistusta, mikä on analogista neuraminidaasi-inhibiittoreille influenssan suhteen.
Lopuksi NORO-virus-inhibiittorin anto saattaa olla yksinkertaista, koska sen on toimittava mahasuolikanavan alueella ”decoy ” mekanismilla, houkuttelemassa virusta. Täten lääkettä ei tarvitse optimoida kehon soluihin otettavaksi tai kuljetettavaksi kehokudoksiin, ja tämä alentaa sivuvaikutuksia muualla kehossa.
http://images.google.se/images?gbv=2&hl=sv&client=firefox-a&rls=org.mozilla%3Aen-US%3Aofficial&sa=3&q=NORO+virus&btnG=S%C3%B6k+bilder
Siis kommenttini: Summa summarum. Ollaan odottamassa uutta pahempaa NORO-virusta Pohjolaan. Mitään rokotetta tai täsmälääkettä ei ole olemassa. Käsienpesu, joka on käytössä pandemista influenssaa estävänä ja muu vastaava hygienia voi auttaa välttämään NORO-virustakin. ( Pesu vedellä ja saippualla ensisijaisesti)
NOROVIRUKSELLA (2009) on uusi variantti etelämailla
Voiko parempi käsihygienia suojata uusilta NORO-virusvarianteilta?
LÄHDE: Kan förbättrad handhygien skydda oss mot nya varianter av norovirus?
LÄHDE: Kjell-Olof Hedlund, Kunskapscentrum för Mikrobiologisk Beredskap, Smittskyddsinstitutet Publicerad i nyhetsbrevet EPI-aktuellt 2009-12-10
Nya norovirus ökar trycket på vården. (Tästä on pitempikin artikkeli olemassa).
En mer aggressiv variant av vinterkräksjukan tycks vara på väg till Sverige. Därför borde sjukhus och institutioner redan nu se över sina städrutiner. Läs hela artikeln i Smittskydd här.
LÄHDE. Publicerad i nyhetsbrevet EPI-aktuellt 2009-12-10
Suomennosta SMIn nettiuutisesta.
LÄHDE: Kan förbättrad handhygien skydda oss mot nya varianter av norovirus?
- (1) Ruotsin maan laboratorioista tulee samantapaista raporttia tavallista vähemmistä NORO-viruspurkauksista ottaen huomioon, mikä vuodenaika nyt on. Tilastolliset numerot ovat vain hieman syksyn tasoa korkeammat NORO-virus valvonnassa. Jopa naapurimaistakin tulee samaa tietoa NORO-viruksen matalammasta aktiivisuudesta hoitolaitoksissa.
- (2) Viime viikkojen matalat raportointinumerot osoittavat samantapaista kehkeytymistä kuin talvisesongin alussa vuonna 2005/2006, jolloin laboratorioista saadut löydöt saavuttivat matalan huippunsa vasta huhtikuussa. Sen kauden matalat numerot voivat selittyä siitä, että ei ollut kiertämässä mitään uutta kantaa NORO-virusgenotyypistä GII.4, mikä on hallinnut hoitoaikoihin liittyneitä mahataudinpurkauksia viime vuosina. Eteläiseltä pallonpuoliskolta talven juuri ohimentyä raportoidaan suuria ongelmia hoitolaitoksien NORO-virusgenotyypin GII.4 vuoden 2008 variantista. Niissä purkauksissa, joita Ruotsin SMI on syksyn aikana analysoinut, ei ole kuitenkaan havaittu tätä eikä muutakaan uutta virusvarianttia.
- (3) Hoitolaitosten ulkopuolista talvisajan mahatautien purkausten valvontaa ei ole, joten SMI:n on vaikea arvioida, onko yhteiskunnassa muuallakin matalaa NORO-virusaktiviteettia parhaillaan. Kuitenkin Tukholman lähistöllä on huhua monen perheen ja esikoulun sairastumisista mahatautiin. Vaikka hoitolaitosten ulkopuolinen yhteiskunta sairastuukin moninaisiin erilaisiin NORO-virustyyppeihin, tapaa NORO-virusgenotyypin GII.4 uudet variantit merkitä vahvaa lisääntymistä mahataudeissa myös yhteiskunnan puolella. SMI esittää toivomuksiaan suhteellisen vähistä talven mahatautipurkauksista vastaisuudessa.
- (4) TARKKA KÄSIEN PESEMINEN on merkityksekästä niitten NORO-virusten leviämisten takia, mitä aina on tapahtumassa, vaikkei olisi uutta viruskantaakaan kyseessä. SMI on saanut päivittäin raportteja vähentyneistä infektioluvuista esim. esikoululaisten keskuudessa ja tämä korreloi parantuneeseen käsihygieniaan, mikä alkoi seurata influenssaepidemian kintereillä. Tieteellistä arviota tästä ei ole vielä olemassa, mutta käsihygienia merkitys on aiemmissakin tutkimuksissakin osoitettu, esim päiväkodeissa on käsihygienia vähentänyt infektioitten rasitetta. Käsihygienian joka asiassa kohennetuilla rutiineilla voinemme olla paremmin varustautuneita siihen hetkeen, kun NORO-viruksen uudet variantit taas alkavat kiertää maassa.
- (5) LUE TARTUNTATAUTIEN INSTITUUTIN TAUTITIEDOTTEISTA RUOTSIKSI NORO-virusta käsittävää: Läs Smittskyddsinstitutets sjukdomsinformation om norovirus här.
LÄHDE: Kjell-Olof Hedlund, Kunskapscentrum för Mikrobiologisk Beredskap, Smittskyddsinstitutet Publicerad i nyhetsbrevet EPI-aktuellt 2009-12-10
- (6) UUSI NORO-virus alkaa lisätä painetta hoitolaitoksissa.
Nya norovirus ökar trycket på vården. (Tästä on pitempikin artikkeli olemassa).
En mer aggressiv variant av vinterkräksjukan tycks vara på väg till Sverige. Därför borde sjukhus och institutioner redan nu se över sina städrutiner. Läs hela artikeln i Smittskydd här.
LÄHDE. Publicerad i nyhetsbrevet EPI-aktuellt 2009-12-10
Suomennosta SMIn nettiuutisesta.
Vanhaa kertausta NORO-viruksista, Caliciviridae
Onpas hankala virus tuo NORO-virus! Otan vanhoista muistiinpanoista aiempaa tietoa, ennenkuin lisään uutta tietoa enemmän.
Siitä saa ruotsinkielistä tietoa lähteestä www.Smittskyddsinstitutet.se
Virus tarttuu hyvin, hyvin tehokkaasti kosketuksen, pisaroiden ja kosketuspintojen välityksellä ja oireet tulevat jo 10-48 tunnissa. Virus on jopa vastustuskykyinen monille desinfektioaineille, joten hygienian lisälläkään se ei oikein ole kontrolloitavaissa. Se voi tarttua myös elintarvikkeen tai veden välityksellä ja aerosolista. Lisäksi se näyttää olevan mitä ihmisrakkain: missä vain ihmisiä vaihtuu paljon tai on paljon koossa. Sairaaloille sellainen on tosi piinallinen tauti.
Lisäksi se antaa vain puolen vuoden immuniteetin ja sitten on taas vaihtanut varianttia. Rokotetta ei ole näköpiirissä. Sairaaloiden osalta joudutaan jopa päivittämään jo muutenkin ylen tarkkoja hygieniasääntöjä ja edelleen keksimään strategioita lisää.
NORO-virusnimen lisäksi sitä sanotaan Norwalk-viruksen kaltaisiin kuuluvaksi virukseksi (NLVs). Sitten sitä sanotaan kaliki-virukseksi, koska se kuuluu sellaiseen virusperheeseen kuin caliciviridae. Sitten lisäksi se ryhmitellään pieniin pyöreisiin selkeän struktuurin omaaviin viruksiin. Joissain internetkuvissa se kyllä näyttää kasvavan kuin sammakonkutu ryväksinä. Yksittäisenä se on on kuin kaunisrusettinen pallo. Sen proteaasista on internetissä kuva. Norovirus ei kykene elämään ihmisen kehon ulkopuolella ja antibiootein siihen ei pysty vaikuttamaan.
KUVA ”NORO virus” fhh.hamburg.de/stadt/Aktuell/behoerden/bsg/hy...
Otan tekstiä englanniksi sen takia, jos joku hakee kuvia Googlen avulla . Saa hakusanoja tästä.
Noroviruses (genus Norovirus, family Caliciviridae) are a group of related, single-stranded RNA, nonenveloped viruses that cause acute gastroenteritis in humans. Norovirus was recently approved as the official genus name for the group of viruses provisionally described as “Norwalk-like viruses” (NLV).
Calici-virus, Virus family Caliciviridae)
small round structured viruses.
Like all viral infections, noroviruses are not affected by treatment with antibiotics, and cannot grow outside of a person’s body.
What are the symptoms of illness caused by noroviruses?
The symptoms of norovirus illness usually include nausea, vomiting, diarrhea, and some stomach cramping. Sometimes people additionally have a low-grade fever, chills, headache, muscle aches, and a general sense of tiredness. The illness often begins suddenly, and the infected person may feel very sick. In most people the illness is self-limiting with symptoms lasting for about 1 or 2 days. In general, children experience more vomiting than adults. Most people with norovirus illness have both of these symptoms.
LÄHDE: http://www.cdc.gov/ncidod/dvrd/revb/gastro/norovirus.htm
Näistä kaikista hankaluuksista akkumuloituu sen ongelmisto ekonomiseksi ja harmittavaksi rasitteeksi, joka voi muun kriisin tullen murtaa yleistä funktionaalista kykyä kriittisissä yhteiskuntaa ylläpitävissä laitoksissa ja tekijöissä. Sitä on vaikea eliminoida ja sen aiheuttama ongelma, dehydraatio (kehon kuivuminen nestehukasta) vaatii rehydraatioterapiaa ( nesteyttävää hoitoa) ja toisaalta sen diagnosointi on vaikeaa, mikä voi hidastaa riittävän rehydraation aloittamisessa.
The Problem:
Noroviruses are the most common cause of gastroenteritis outbreaks and are estimated to cause 23 million cases in the United States alone each year. Norovirus infection occurs worldwide, and it is a frequent cause of traveler’s diarrhea. Caliciviruses are classified as Category B bioterrorism agents because they are very contagious and can be easily transmitted by water, food, aerosols, and by person-to-person spread; require only a very low dose to cause an infection; are highly stable in the environment.
Large outbreaks of the disease can cause economic problems and could potentially overwhelm the capabilities of public health, hospital, and military facilities. Furthermore, once a facility becomes contaminated, it is difficult and expensive to eliminate the virus. There is currently no way to treat calicivirus infections, other than rehydration therapy. Calicivirus infection is also difficult to diagnose. There is no vaccine to prevent calicivirus infection at this time.
http://www.bcm.edu/molvir/eidbt/images/noro1.jpg
www.bcm.edu/molvir/eidbt/eidbt-mvm-noro.htm
LÄHDE: Rockx BH et al. Association of histo-blood group antigens and susceptibility to norovirus infections. J Infect Dis. 2005 Mar 1;191(5):749-54. Epub 2005 Jan 25.
BACKGROUND: Noroviruses (NoVs) are the leading cause of viral gastroenteritis in humans of all ages. Challenge studies that used the NoV prototype strain Norwalk virus (NV) have shown that some individuals are not susceptible to infection, suggesting the absence of a receptor.
Recent studies have identified histo-blood group antigens (HBGAs) as Being a possible receptors.nonsecretor and presence of HBGA type B were associated with protection against infection with NV, a genogroup (GG) I NoV.
CONCLUSIONS: Individuals with the HBGA type B may be protected against infection with GGI (but not GGII) NoVs. The association between susceptibility to NoV infection and being a secretor may be restricted to GGI NoV.
LÄHDE: Larsson MM et al. Antibody prevalence and titer to norovirus (genogroup II) correlate with secretor (FUT2) but not with ABO phenotype or Lewis (FUT3) genotype. Division of Molecular Virology, University of Linkoping, 581 85 Linkoping, Sweden.
BACKGROUND: Histo-blood group antigens and secretor status have been associated with susceptibility to Norovirus infections, which suggests that antibody prevalence and titer might correlate with these phenotypes.
METHODS: Plasma samples (n = 105) from Swedish blood donors that had been genotyped for secretor (FUT2) and Lewis (Le; FUT3) genotypes and phenotyped for ABO and Le blood groups, were analyzed for immunoglobulin G antibody prevalence and titers to norovirus genogroup (GG) II.4.
RESULTS: The results showed that nonsecretors (se4128se428) and Le a+b- individuals not only had significantly lower antibody titers than did secretors (P < .0001) and Le a-b+ individuals (P < .0002) but were also significantly more often antibody negative (P < .05). Antibody titers in secretors were not significantly different between individuals of different Le (FUT3) genotypes or different ABO phenotypes. CONCLUSIONS: Nonsecretors and Le a+b- individuals are significantly less prone to be infected with GGII noroviruses. This new information extends previous knowledge and supports the hypothesis that nonsecretors are relatively but not absolutely resistant to norovirus infections.
Ruotsissa kerrotaan olevan vaivaa seuraavista genotyypeistä, jotka vaikuttavat olevan samat kuin Suomen virustyypit:
”Vi befinner oss mitt i vinterns norovirusepidemi och laboratorierna rapporterar fortfarande många virusfynd. Aktuella typningsresultat från december och januari visar att samtliga norovirusstammar från 25 vårdrelaterade utbrott tillhör genotyp G2.4 variant 2006 a och b (definierade enligt FBVE Norovirus Quick Typing Database.) Det är med andra ord samma stamvarianter som debuterade Sjukdomsinformation om calicivirus (noro- och sapovirus)”.
Muistiin päivitystä NORO-viruksesta vuodelta 2007.
Tämä on virus, josta varmaan saa kuulla uutisia vähän väliä. Taksonomiasta
(taxonomy) voi katsoa sitä isoa RNA virus joukkoa, mihin tämä Caliciviridae asettuu yhtenä erilliseksi merkattuna, vähemmän tunnettuna siis aika uudehkona virusryhmänä. Vaikka oikeastaan kaikki mikä koskee nukliinihappotiedettä (DNA tai RNA), ei ole mitään kovin vanhaa.
”WIKIPEDIA Group IV - positive-sense ssRNA viruses”
Family Caliciviridae
2007-03-05 19:09
2007-12-05 22:24 Nokialla NOROVIRUSTA. Armeija auttaa, mutta voi itse saada tartunnan. Nokian vesikatastrofi ja mahataudit olivat tosi paha asia.
- 2007-03-05 19:39 Lääkärilehden pääkirjoitukseen oli asetettu NORO-virus ”Norovirus sairaaloiden kiusana” SLL numero 9; 2007. Sivu 865. Saksassakin oli silloin raportoitu 600 epidemiaa. Yksi ainoa oksennus pystyisi tartuttamaan koko Suomen kansan, kertoo sen päivän Suomen Lääkärilehti.
- Mikäs se tällainen ihmeen hankala virus oikein on?
Siitä saa ruotsinkielistä tietoa lähteestä www.Smittskyddsinstitutet.se
Virus tarttuu hyvin, hyvin tehokkaasti kosketuksen, pisaroiden ja kosketuspintojen välityksellä ja oireet tulevat jo 10-48 tunnissa. Virus on jopa vastustuskykyinen monille desinfektioaineille, joten hygienian lisälläkään se ei oikein ole kontrolloitavaissa. Se voi tarttua myös elintarvikkeen tai veden välityksellä ja aerosolista. Lisäksi se näyttää olevan mitä ihmisrakkain: missä vain ihmisiä vaihtuu paljon tai on paljon koossa. Sairaaloille sellainen on tosi piinallinen tauti.
Lisäksi se antaa vain puolen vuoden immuniteetin ja sitten on taas vaihtanut varianttia. Rokotetta ei ole näköpiirissä. Sairaaloiden osalta joudutaan jopa päivittämään jo muutenkin ylen tarkkoja hygieniasääntöjä ja edelleen keksimään strategioita lisää.
- Katsotaan vähän lisää NORO virustietoa netistäkin. Hakusanana voi pitää sen sukunimeä. ”Caliciviruses, NORO-virus”
NORO-virusnimen lisäksi sitä sanotaan Norwalk-viruksen kaltaisiin kuuluvaksi virukseksi (NLVs). Sitten sitä sanotaan kaliki-virukseksi, koska se kuuluu sellaiseen virusperheeseen kuin caliciviridae. Sitten lisäksi se ryhmitellään pieniin pyöreisiin selkeän struktuurin omaaviin viruksiin. Joissain internetkuvissa se kyllä näyttää kasvavan kuin sammakonkutu ryväksinä. Yksittäisenä se on on kuin kaunisrusettinen pallo. Sen proteaasista on internetissä kuva. Norovirus ei kykene elämään ihmisen kehon ulkopuolella ja antibiootein siihen ei pysty vaikuttamaan.
KUVA ”NORO virus” fhh.hamburg.de/stadt/Aktuell/behoerden/bsg/hy...
Otan tekstiä englanniksi sen takia, jos joku hakee kuvia Googlen avulla . Saa hakusanoja tästä.
Noroviruses (genus Norovirus, family Caliciviridae) are a group of related, single-stranded RNA, nonenveloped viruses that cause acute gastroenteritis in humans. Norovirus was recently approved as the official genus name for the group of viruses provisionally described as “Norwalk-like viruses” (NLV).
Calici-virus, Virus family Caliciviridae)
small round structured viruses.
Like all viral infections, noroviruses are not affected by treatment with antibiotics, and cannot grow outside of a person’s body.
- Mitä kaikkea NORO-virus voi aiheuttaa?
What are the symptoms of illness caused by noroviruses?
The symptoms of norovirus illness usually include nausea, vomiting, diarrhea, and some stomach cramping. Sometimes people additionally have a low-grade fever, chills, headache, muscle aches, and a general sense of tiredness. The illness often begins suddenly, and the infected person may feel very sick. In most people the illness is self-limiting with symptoms lasting for about 1 or 2 days. In general, children experience more vomiting than adults. Most people with norovirus illness have both of these symptoms.
LÄHDE: http://www.cdc.gov/ncidod/dvrd/revb/gastro/norovirus.htm
- Mikä sen ongelma on, jos se menee tautina noin nopeasti ohi?
Näistä kaikista hankaluuksista akkumuloituu sen ongelmisto ekonomiseksi ja harmittavaksi rasitteeksi, joka voi muun kriisin tullen murtaa yleistä funktionaalista kykyä kriittisissä yhteiskuntaa ylläpitävissä laitoksissa ja tekijöissä. Sitä on vaikea eliminoida ja sen aiheuttama ongelma, dehydraatio (kehon kuivuminen nestehukasta) vaatii rehydraatioterapiaa ( nesteyttävää hoitoa) ja toisaalta sen diagnosointi on vaikeaa, mikä voi hidastaa riittävän rehydraation aloittamisessa.
The Problem:
Noroviruses are the most common cause of gastroenteritis outbreaks and are estimated to cause 23 million cases in the United States alone each year. Norovirus infection occurs worldwide, and it is a frequent cause of traveler’s diarrhea. Caliciviruses are classified as Category B bioterrorism agents because they are very contagious and can be easily transmitted by water, food, aerosols, and by person-to-person spread; require only a very low dose to cause an infection; are highly stable in the environment.
Large outbreaks of the disease can cause economic problems and could potentially overwhelm the capabilities of public health, hospital, and military facilities. Furthermore, once a facility becomes contaminated, it is difficult and expensive to eliminate the virus. There is currently no way to treat calicivirus infections, other than rehydration therapy. Calicivirus infection is also difficult to diagnose. There is no vaccine to prevent calicivirus infection at this time.
http://www.bcm.edu/molvir/eidbt/images/noro1.jpg
www.bcm.edu/molvir/eidbt/eidbt-mvm-noro.htm
- Kun katsoo internetmaisemaa NORO-viruskuvista, huomaa että tärkeimpänä ohjeena korostetaan kaikenlaista hygieniaa.
- NORO-virus on tieteen pihdeissä!
LÄHDE: Rockx BH et al. Association of histo-blood group antigens and susceptibility to norovirus infections. J Infect Dis. 2005 Mar 1;191(5):749-54. Epub 2005 Jan 25.
BACKGROUND: Noroviruses (NoVs) are the leading cause of viral gastroenteritis in humans of all ages. Challenge studies that used the NoV prototype strain Norwalk virus (NV) have shown that some individuals are not susceptible to infection, suggesting the absence of a receptor.
Recent studies have identified histo-blood group antigens (HBGAs) as Being a possible receptors.nonsecretor and presence of HBGA type B were associated with protection against infection with NV, a genogroup (GG) I NoV.
CONCLUSIONS: Individuals with the HBGA type B may be protected against infection with GGI (but not GGII) NoVs. The association between susceptibility to NoV infection and being a secretor may be restricted to GGI NoV.
- Linköpingissä on NORO-virus ollut tieteen fokuksessa.
LÄHDE: Larsson MM et al. Antibody prevalence and titer to norovirus (genogroup II) correlate with secretor (FUT2) but not with ABO phenotype or Lewis (FUT3) genotype. Division of Molecular Virology, University of Linkoping, 581 85 Linkoping, Sweden.
BACKGROUND: Histo-blood group antigens and secretor status have been associated with susceptibility to Norovirus infections, which suggests that antibody prevalence and titer might correlate with these phenotypes.
METHODS: Plasma samples (n = 105) from Swedish blood donors that had been genotyped for secretor (FUT2) and Lewis (Le; FUT3) genotypes and phenotyped for ABO and Le blood groups, were analyzed for immunoglobulin G antibody prevalence and titers to norovirus genogroup (GG) II.4.
RESULTS: The results showed that nonsecretors (se4128se428) and Le a+b- individuals not only had significantly lower antibody titers than did secretors (P < .0001) and Le a-b+ individuals (P < .0002) but were also significantly more often antibody negative (P < .05). Antibody titers in secretors were not significantly different between individuals of different Le (FUT3) genotypes or different ABO phenotypes. CONCLUSIONS: Nonsecretors and Le a+b- individuals are significantly less prone to be infected with GGII noroviruses. This new information extends previous knowledge and supports the hypothesis that nonsecretors are relatively but not absolutely resistant to norovirus infections.
- Mikä NORO-virustyyppi on raivoamassa Pohjoismaissa?
Ruotsissa kerrotaan olevan vaivaa seuraavista genotyypeistä, jotka vaikuttavat olevan samat kuin Suomen virustyypit:
”Vi befinner oss mitt i vinterns norovirusepidemi och laboratorierna rapporterar fortfarande många virusfynd. Aktuella typningsresultat från december och januari visar att samtliga norovirusstammar från 25 vårdrelaterade utbrott tillhör genotyp G2.4 variant 2006 a och b (definierade enligt FBVE Norovirus Quick Typing Database.) Det är med andra ord samma stamvarianter som debuterade Sjukdomsinformation om calicivirus (noro- och sapovirus)”.
- Kaikesta päätellen asiaan on kuitenkin tulemassa tieteen taholta lisä valoa lähivuosina.
Muistiin päivitystä NORO-viruksesta vuodelta 2007.
Tämä on virus, josta varmaan saa kuulla uutisia vähän väliä. Taksonomiasta
(taxonomy) voi katsoa sitä isoa RNA virus joukkoa, mihin tämä Caliciviridae asettuu yhtenä erilliseksi merkattuna, vähemmän tunnettuna siis aika uudehkona virusryhmänä. Vaikka oikeastaan kaikki mikä koskee nukliinihappotiedettä (DNA tai RNA), ei ole mitään kovin vanhaa.
”WIKIPEDIA Group IV - positive-sense ssRNA viruses”
Family Caliciviridae
2007-03-05 19:09
2007-12-05 22:24 Nokialla NOROVIRUSTA. Armeija auttaa, mutta voi itse saada tartunnan. Nokian vesikatastrofi ja mahataudit olivat tosi paha asia.
NOROVIRUS ja antiadheesioterapia
Uusi väitöskirja NORO-viruksesta
LÄHDE: RYDELL Gustav E. Norovirus, causative agent of wintervomiting disease, exploits several histo-blood group glycans for adhesion. University of Gothenburg 2009.
ISBN978-91-628-7794-1
Abstraktin suomennosta.
NOROVIRUS tunnetaan kautta maailman gastroenteriittipurkausten aikaansaajana, mutta sitä vastaan ei kuitenkaan ole rokotteita tai lääkkeitä, jolla tätä virustautia voisi estää tai hoitaa. Kun on tehty virusten kaltaisten partikkeleitten (VLPs) avulla altistustutkimuksia tai sitoutumistutkimuksia, on saatu viitteitä norovirukselle alttiuden korreloivan henkilön sekretoriseen statukseen ( secretor status).
Tämä väitöskirja antaa tukea ajatukselle osoittamalla, että 105:n ruotsalaisen verenluovuttajan joukossa non-sekretorista tyyppiä edustavilla henkilöillä oli merkitsevästi pienemmeät tiitterit norovirusgenotyypille(G) II.4 spesifisiä IgG ryhmän vasta-aineita sekretorisiin yksilöihin verrattuna. Kuitenkin muutamilla non-sekretorisilla yksilöillä oli korkeat vasta-ainetiitterit, mikä antaisi viitettä siitä, että on olemassa myös sekretorisesta statuksesta riippumattomia viruskantoja.
Koska puuttuu in vitro-viljely metodeita, käytettiin VLP:itä, virusten kaltaisia partikkeleita, kun luonnehdittiin glykaaniin sitoutumisominaisuuksia eri noroviruskannoista. Sellaiset VLP:t kuten Chron1(GII.3) ja Dijon (GII.4) viruskannat tunnistivat sylkinäytteitä sekretorisista yksilöistä, mutta eivät non-sekretorisista. Kun käytettiin neoglykoproteiineja pystyttiin osoittamaan näiden kahden VLP:n tunnistavan sekretorisesta geenistä riippuvan glykaanin lisäksi seuraavia rakenteita: sialyl Lewis x-epitooppia, sialyl diLewis x:n rakenteellisia analogeja ja sialyloitua tyyppi 2:ta. Mutta sitävastoin Norwalk viruskannan(GI.1) VLP:t tunnistivat vain sekretorisesta geenistä riippuvaa glykaania.
Inhibitiokokeissa (estämiskokeissa) sialyyli Lewis x - konjugaatti pystyi täydellisesti blokeeraamaan Chron1 ja Dijon VLP:n sitoutumisen sylkinäytteisiin.
Etsittäessä reseptoriglykokonjugaatteja osoitettiin tällä työllä nyt ensimmäisen kerran, että ihmisen noroviruksen VLP:t pystyvät sitomaan glykosfingolipidiä.
Tutkija käytti kromatogrammi-sitoutumistekniikkaa ja radioaktiivisesti merkattuja Norwalk VLP:tä ja osoitti sen tunnistavan veriryhmien A ja H mutta ei B-veriryhmän epitooppeja. VLP-sitoutuminen glykosfingolipideihin lipidikaksoiskerroksessa monitoroitiin QCM-D tekniikalla ( Quartz crystal microbalance with dissipation). Norwalk ja Dijon VLP sitoutuivat H tyyppiä 1 sisältäviin kaksoiskerroksiin, mutta eivät sitoutuneet niihin glykosfingolipideihin, joissa oli Lewis a epitooppi. Multivalenttisuuttta osoittaa , että molemmat VLP:t näyttivät omaavan jonkin kynnyspitoisuuden H tyyppiä 1 kohtaan, ja kynnyksen alittavissa pitoisuuksissa ei tapahtunut sitoutumista.
Johtopäätös. Tämä väitöskirja kuvaa monenlaisia kudos-veriryhmäglykokonjugaatteja, joita ihmisen norovirukset tunnistavat ja tämä antaa viitettä uusista lähestymistavoista, kun ollaan hahmottamassa glykomimeettejä (glycomimetics) noroviruksen vastaiseen anti-adheesioterapiaan.
tisdag 1 december 2009
Uusia antivirusstrategioita tulevaisuudessa
LÄHDE:
Gong J, Fang H, Li M, Liu Y, Yang K, Liu Y, Xu W. Potential targets and their relevant inhibitors in anti-influenza fields. Curr Med Chem. 2009;16(28):3716-39. Department of Medicinal Chemistry, School of Pharmaceutical Sciences, Shandong University, Jinan, Shandong, China.
INFLUENSSA on tauti, joka iskee vahvasti miljooniin ihmisiin vuosittain Äskettäin on jälleen pohdittu patogeenisen A H5N1 lintuinfluenssaviruksen merkitystä ja sen mahdollista pandemista potentiaalia ihmiskunnassakin.
Sitä myöten kun viruksia koskeva biologia kehittyy, löytyy yhä useampia antivirusstrategioita näitä viruksia kohtaan.
Otsikon artikkelissa tehdään katsausta, jossa pohditaan useita jo aiemmin kokeiltuja sekä mahdollisia anti-influenssalääkekohteita, joihin kuuluu
Gong J, Fang H, Li M, Liu Y, Yang K, Liu Y, Xu W. Potential targets and their relevant inhibitors in anti-influenza fields. Curr Med Chem. 2009;16(28):3716-39. Department of Medicinal Chemistry, School of Pharmaceutical Sciences, Shandong University, Jinan, Shandong, China.
INFLUENSSA on tauti, joka iskee vahvasti miljooniin ihmisiin vuosittain Äskettäin on jälleen pohdittu patogeenisen A H5N1 lintuinfluenssaviruksen merkitystä ja sen mahdollista pandemista potentiaalia ihmiskunnassakin.
Sitä myöten kun viruksia koskeva biologia kehittyy, löytyy yhä useampia antivirusstrategioita näitä viruksia kohtaan.
Otsikon artikkelissa tehdään katsausta, jossa pohditaan useita jo aiemmin kokeiltuja sekä mahdollisia anti-influenssalääkekohteita, joihin kuuluu
- virustekijöitä kuten HA, hemagglutiniini, M2 jonikanavaproteiini, RNA:sta riippuva RNA-polymeraasi RdRp, nukleoproteiini NP, nonstrukturelli proteiini NS ja neuraminidaasi NA
- isäntäkehoon kuuluvia tekijöitä ( kuten v-ATPaasi, inosiinimonofosfaattidehydrogenaasi, IMPDH ja intrasellulaariset signaalikaskadit )
- näiden relevantteja estäjiä eli inhibiittoreita.
NLS1 ja NLS2 AH1N1 NP proteiinissa.
http://www.virologyj.com/content/4/1/49Sitaatti (engl), jonka sisältönä on selvitysNP-proteiinin NLS 1 ja NLS 2 kohtien tärkeydestä virusgenomin siirtämisessä isäntäsolun tumaan kopioitumaan ja lisääntymään.
Sitaatti (engl), jonka sisältönä on selvitysNP-proteiinin NLS 1 ja NLS 2 kohtien tärkeydestä virusgenomin siirtämisessä isäntäsolun tumaan kopioitumaan ja lisääntymään.
Discussion
Using influenza vRNPs purified under acidic conditions, and thus mimicking physiologically-relevant influenza infections as closely as possible, we have found that inhibition of either NLS1 or NLS2 on influenza NP significantly inhibited the nuclear import of influenza vRNP complexes. Therefore, both NLS1 and NLS2 on NP are involved in mediating the nuclear import of incoming vRNP complexes. These two sequences act independently of each other, as peptide competition with or antibody inhibition of only one of these sequences still resulted in a certain, though less pronounced, degree of nuclear import of vRNPs. Furthermore, when both NLS1 and NLS2 were competed with peptides or blocked by antibodies, the nuclear import of the vRNPs was even more drastically reduced.
Some differences, however, existed in the ability of the NLS peptides versus the anti-NLS antibodies to inhibit nuclear import of the vRNPs. These differences were likely due to the nature of the competition experiments, since peptide competition with the vRNPs for the cytosolic nuclear import receptors is less specific than direct inhibition of the vRNP NLSs with antibodies. The antibody inhibition experiments may hence provide a more accurate picture of the relative contributions of NLS1 and NLS2 to influenza vRNP nuclear import. From these results, it appears that peptides mimicking or antibodies against these conserved NLS regions on NP may be an effective means of interrupting a critical stage in the influenza A life cycle.
Interestingly, performing a sequence alignment (using Clustal W [43]) of NP from different influenza A strains, we can observe that each of NLS1 and NLS2 on NP are highly conserved among influenza A strains. However, NLS1 and NLS2 do not share much similarity with any region on NP of influenza B or C, as analyzed from a Clustal W alignment. This is in agreement with previous studies that have not been able to pinpoint the NLS in NP for influenza B [44], so it appears that influenza B and C may utilize NLSs that are different from those of influenza A.
With respect to NLS1, the only residue conserved among influenza A, B, and C is a conserved arginine at position 8 of the influenza A NP, which agrees with previous studies indicating that that residue is one of the most important residues involved in mediating the nuclear import of recombinant NP .
For NLS2, residue 214 on influenza A NP is probably one of the most important residues as an arginine or lysine is found in that position in influenza A, B, and C.
The exact length of NLS2 also remains to be determined. Previous studies had implicated that NLS2 was a bipartite NLS of 19 amino acids long, spanning residues 198 to 216 [31]. However, recent structural data has questioned that this NLS functions as a bipartite classical NLS since the crystal structure of NP showed that the two clusters of basic residues of the bipartite NLS2 on NP were located too close together in space to be functional as a bipartite NLS [5]. Even though NLS2 may not be a bipartite NLS, the relevance of residues 213, 214, and 216 on NP in mediating the nuclear import of recombinant NP appears to be significant [31]. However, which other residues in NLS2 are important in mediating influenza nuclear import remains to be determined.
Our studies here with influenza vRNPs also confirm findings with recombinant NP that NLS1 is the stronger of the two NLSs [29,31,37]. From the crystal structure of NP [5], it is probably reasonable to assume that NLS1, being an N-terminal sequence near the edge of NP, may be more accessible to the binding of cytosolic nuclear import factors than NLS2.
However, having both NLS1 and NLS2 as functional NLSs on influenza vRNPs could serve a vital purpose to the virus. If, in the event that the N-terminal NLS1 is inadvertently cleaved off by any proteases in its host cell, the vRNP may still have an extra NLS (NLS2) to mediate its nuclear import. Nonetheless, it appears that with so many copies of NP, it does seem to be a redundant function.
However, it is not clear how many of these NLSs are exposed when NP oligomerizes and associates with the vRNA. Therefore, further studies at understanding the kinetics, cellular targets, conformational states, and role in viral replication of NLS1 and NLS2 will be required to bring further light to their roles in influenza cellular trafficking and replication.
Previous work by other groups have concentrated mainly on the nuclear import of recombinant NP [5,27-31,37]. These studies have provided a better understanding of the role of the various NLSs on NP in the nuclear import of newly-synthesized NP, which occurs after the initial nuclear import of the entire vRNP complex and the subsequent synthesis of new NP in the cytoplasm [12,14]. To study the nuclear import of influenza vRNPs, O'Neil et al. (1995) [27], and more recently Cros et al. (2005) [26], formed in vitro-assembled NP-RNA complexes by incubating recombinant NP with in vitro-synthesized influenza vRNA. To study the nuclear import of the influenza genome, however, vRNPs purified from influenza virions would likely be the preferred substrates over in vitro-formed vRNA-NP complexes. This is because the actual assembly of NP into an oligomeric structure, and the interactions of NP with the vRNA in actual influenza infections, would likely result in structural differences between in vitro-formed RNA-NP complexes versus virally-produced and assembled vRNPs. For example, certain NLSs may be exposed or hidden according to how NP actually interacts with itself in the oligomer and how NP interacts with the vRNA. In addition, NP molecules within actual influenza vRNPs that are produced in mammalian cells may have differences in their post-translational modifications compared to recombinant NP molecules produced in bacteria. Furthermore, any conformational changes in the structure of the vRNPs after influenza export from the cell, viral entry into a new cell, and during or after their exit from endosomes are not taken into account by in vitro-formed RNA-NP complexes. The methodological differences in the preparation of vRNPs may therefore explain the differences observed in studies using in vitro-formed RNP and our results reported here using naturally-occurring, influenza-derived vRNP complexes. For example, Cros et al. [27] found that disruption of NLS2 on NP has no effect on the nuclear accumulation of in vitro-formed vRNA-NP complexes, while we showed here that interfering with NLS2 on NP diminished the nuclear import capability of influenza-purified vRNPs (Figs. 4 and 5). Likewise, Ozawa et al. [37] found that NLS2, but not NLS1, deletion mutants of NP were unable to target effectively to nucleolar regions, while we showed here that nucleolar localization still occurred whether NLS1 or NLS2 on NP was inhibited.
Much work on understanding the nuclear import of the influenza genome still remains. For example, the nuclear accumulation sequence (NAS) within influenza-assembled vRNP complexes is still a mystery. The NAS on NP was originally found to mediate nuclear accumulation of NP in Xenopus oocytes [45], but more recently has been found to be a cytoplasmic retention signal in mammalian cells [30,31,46]. It would therefore be useful to understand in greater detail the role of the NAS and how its function relates to that of NLS1 and NLS2.
In addition, the role of M1 in preventing nuclear import of vRNP is still unclear [11,39]. For example, M1 may be acting indirectly, where interaction of M1 with the vRNPs causes the vRNPs to change their structural conformation [47] and thus expose or hide certain NLSs. Alternatively, M1 may be binding directly to the NLSs of vRNPs to inhibit their nuclear import.
The studies completed to date also raise the question as to under what conditions and in what conformational states of NP would NLS1 and NLS2 act to mediate the nuclear import of vRNPs. Further unraveling the answers to these questions may give us a more detailed understanding of how the various NLSs on the influenza vRNPs work together or independently to mediate influenza A nuclear import.
Conclusion
In summary, we have showed in this study that inhibition of either NLS1 or NLS2 on NP from influenza-derived vRNP complexes significantly decreased the extent of nuclear localization of the influenza genome.
Furthermore, inhibiting both NLSs resulted in an additive effect, causing an even greater decrease in vRNP nuclear accumulation.
This indicates that both NLS1 and NLS2 on NP play a critical role in nuclear import by acting independently to mediate the nuclear import of incoming influenza A vRNP complexes.
The importance of our findings in the design and development of novel influenza antiviral therapeutics is critical, as both NLSs will likely require to be inhibited to more completely abolish influenza nuclear import and thus viral replication.
Nukleokapsidiproteiini NP ja Geeni 5.
NUKLEOKAPSIDIPROTEINI Geeni 5 tuote.
Norjalaisen viruksen NP netistä. Vertaan Meksikossa ilmenneesseen keväiseen virukseen.
/organism="Influenza A virus (A/Norway/3206-3/2009(H1N1))"
/strain="A/Norway/3206-3/2009(H1N1)"
/serotype="H1N1"
/isolation_source="gender:M; age:adult; post mortem lung
http://www.ncbi.nlm.nih.gov/protein/269978856
/country="Norway: Oslo"
/collection_date="01-Sep-2009"
Protein 1..498
/product="nucleocapsid protein"
/gene="NP"
/coded_by="CY052020.1:18..1514"
ORIGIN
1 masqgtkrsy eqmetdgerq dateirasvg rmiggigrfy iqmctelkls dydgrliqns
61 itiermvlsa fderrnkyle ehpsagkdpk ktggpiyrri dgkwmrelil ydkeeirrvw
121 rqanngedat aglthimiwh snlndatyqr tralvrtgmd prmcslmqgs tlprrsgaag
181 aavkgvgtia melirmikrg indrnfwrge ngrrtrvaye rmcnilkgkf qtaaqrammd
241 qvresrnpgn aeiedlifla rsalilrgsv ahksclpacv yglavasghd feregyslvg
301 idpfkllqns qvvslmrpne npahksqlvw machsaafed lrvssfirgk kviprgklst
361 rgvqiasnen vetmdsntle lrsrywairt rsggntnqqk asagqisvqp tfsvqrnlpf
421 eratvmaafs gnnegrtsdm rtevirmmes akpedlsfqg rgvfelsdek atnpivpsfd
481 msnegsyffg dnaeeyds
ALLA: (A/California/04/2009(H1N1)
/collection_date="01-Apr-2009"
Protein 1..498
ORIGIN
1 masqgtkrsy eqmetggerq dateirasvg rmiggigrfy iqmctelkls dydgrliqns
61 itiermvlsa fderrnkyle ehpsagkdpk ktggpiyrrv dgkwmrelil ydkeeirrvw
121 rqanngedat aglthimiwh snlndatyqr tralvrtgmd prmcslmqgs tlprrsgaag
181 aavkgvgtia melirmikrg indrnfwrge ngrrtrvaye rmcnilkgkf qtaaqrammd
241 qvresrnpgn aeiedlifla rsalilrgsv ahksclpacv yglavasghd feregyslvg
301 idpfkllqns qvvslmrpne npahksqlvw machsaafed lrvssfirgk kviprgklst
361 rgvqiasnen vetmdsntle lrsrywairt rsggntnqqk asagqisvqp tfsvqrnlpf
421 eratvmaafs gnnegrtsdm rtevirmmes akpedlsfqg rgvfelsdek atnpivpsfd
481 msnegsyffg dnaeeyds
KOMMENTINI:
Löydän vain kaksi aminohappomuutosta näiden kesken. .
Kalifornian G16 ja V200 on norjalaisessa D ja i(isoleusiini)
G 16 D ja V 100 (I).
Siis nukleokapsidiproteiini NP vaikuttaa olevan aika stabiili.
Koskeeko muutos NLS1 tai NLS2 kohtaa, jotka määräävät tumaan pääsynopeuden?
NLS1 kohdassa on tärkeä R8, arginiini asemassa 8. Sekä norjalaisessa että keväisessä viruksessa on 8Arg.
Mitä NLS2 kohtaan tulee: Aminohappo 214 on ehkä NP:ssa eräs kaikkein tärkein aminohappotähde . A, B ja C-influenssavirusten NP proteiineissa on kaikissa siinä joko R(arginiini) tai K(lysiini).
Norjalaisessa ja kevään kalifornialaisessa AH1N1 viruksessa on NP-proteiinissa R214 molemmilla viruksilla, joten tumaanpääsy lie hyvä.
NLS2 lienee haettava 198-216 jaksolta.krg indrnfwrge ngrrtr, tämä on molemmissa sama.
Norjalaisen viruksen NP netistä. Vertaan Meksikossa ilmenneesseen keväiseen virukseen.
/organism="Influenza A virus (A/Norway/3206-3/2009(H1N1))"
/strain="A/Norway/3206-3/2009(H1N1)"
/serotype="H1N1"
/isolation_source="gender:M; age:adult; post mortem lung
http://www.ncbi.nlm.nih.gov/protein/269978856
/country="Norway: Oslo"
/collection_date="01-Sep-2009"
Protein 1..498
/product="nucleocapsid protein"
/gene="NP"
/coded_by="CY052020.1:18..1514"
ORIGIN
1 masqgtkrsy eqmetdgerq dateirasvg rmiggigrfy iqmctelkls dydgrliqns
61 itiermvlsa fderrnkyle ehpsagkdpk ktggpiyrri dgkwmrelil ydkeeirrvw
121 rqanngedat aglthimiwh snlndatyqr tralvrtgmd prmcslmqgs tlprrsgaag
181 aavkgvgtia melirmikrg indrnfwrge ngrrtrvaye rmcnilkgkf qtaaqrammd
241 qvresrnpgn aeiedlifla rsalilrgsv ahksclpacv yglavasghd feregyslvg
301 idpfkllqns qvvslmrpne npahksqlvw machsaafed lrvssfirgk kviprgklst
361 rgvqiasnen vetmdsntle lrsrywairt rsggntnqqk asagqisvqp tfsvqrnlpf
421 eratvmaafs gnnegrtsdm rtevirmmes akpedlsfqg rgvfelsdek atnpivpsfd
481 msnegsyffg dnaeeyds
ALLA: (A/California/04/2009(H1N1)
/collection_date="01-Apr-2009"
Protein 1..498
ORIGIN
1 masqgtkrsy eqmetggerq dateirasvg rmiggigrfy iqmctelkls dydgrliqns
61 itiermvlsa fderrnkyle ehpsagkdpk ktggpiyrrv dgkwmrelil ydkeeirrvw
121 rqanngedat aglthimiwh snlndatyqr tralvrtgmd prmcslmqgs tlprrsgaag
181 aavkgvgtia melirmikrg indrnfwrge ngrrtrvaye rmcnilkgkf qtaaqrammd
241 qvresrnpgn aeiedlifla rsalilrgsv ahksclpacv yglavasghd feregyslvg
301 idpfkllqns qvvslmrpne npahksqlvw machsaafed lrvssfirgk kviprgklst
361 rgvqiasnen vetmdsntle lrsrywairt rsggntnqqk asagqisvqp tfsvqrnlpf
421 eratvmaafs gnnegrtsdm rtevirmmes akpedlsfqg rgvfelsdek atnpivpsfd
481 msnegsyffg dnaeeyds
KOMMENTINI:
Löydän vain kaksi aminohappomuutosta näiden kesken. .
Kalifornian G16 ja V200 on norjalaisessa D ja i(isoleusiini)
G 16 D ja V 100 (I).
Siis nukleokapsidiproteiini NP vaikuttaa olevan aika stabiili.
Koskeeko muutos NLS1 tai NLS2 kohtaa, jotka määräävät tumaan pääsynopeuden?
NLS1 kohdassa on tärkeä R8, arginiini asemassa 8. Sekä norjalaisessa että keväisessä viruksessa on 8Arg.
Mitä NLS2 kohtaan tulee: Aminohappo 214 on ehkä NP:ssa eräs kaikkein tärkein aminohappotähde . A, B ja C-influenssavirusten NP proteiineissa on kaikissa siinä joko R(arginiini) tai K(lysiini).
Norjalaisessa ja kevään kalifornialaisessa AH1N1 viruksessa on NP-proteiinissa R214 molemmilla viruksilla, joten tumaanpääsy lie hyvä.
NLS2 lienee haettava 198-216 jaksolta.krg indrnfwrge ngrrtr, tämä on molemmissa sama.
måndag 30 november 2009
Matrixproteiini M2 muuntelee.
MATRIX PROTEIINI M1, M2.
Geeni 7 tuote.
Norjalaisen viruksen M1 proteiini: Sen C-terminaalinen jakso( samanlainen kuin esim katalonialaisen)
/collection_date="08-Sep-2009"
Protein 1..252
/product="matrix protein 1"
Region 2..157
/region_name="Flu_M1"
/note="Influenza Matrix protein (M1); pfam00598"
/db_xref="CDD:109646"
Region 158..252
/region_name="Flu_M1_C"
/note="Influenza Matrix protein (M1) C-terminal domain;
smart00759"
/db_xref="CDD:128998"
CDS 1..252
/gene="M1"
/coded_by="CY052001.1:11..769"
ORIGIN
1 mslltevety vlsiipsgpl kaeiaqrles vfagkntdle almewlktrp ilspltkgil
61 gfvftltvps erglqrrrfv qnalngngdp nnmdravkly kklkreitfh gakevslsys
121 tgalascmgl iynrmgtvtt eaafglvcat ceqiadsqhr shrq//matttn plirhenrmv
181 lasttakame qmagsseqaa eamevanqtr qmvhamrtig thpsssaglk ddllenlqay
241 qkrmgvqmqr fk
Norjalaisen viruksen M2 proteiinista annetaan kaksi hieman poikkeavaa versiota.
//
10 20 30 40 50 60 70 80
....*....|....*....|....*....|....*....|....*....|....*....|....*....|....*....|
gi 81936884 1 MSLLTEVETPIKSGWECRCNDSSDLLVAIASITGILHLILWIFDRLFFKCAYRRFRHGLKRGPSTGGIPESMREEYRQEQ 80
gi 82053560 1 MSLLTEVETPTRNEWECRCSDSSDPLVVAASIIGILHLILWILDRLFFKCIYRRLKYGLKRGPATAGVPESMREEYRQEQ 80
90
....*....|....*..
gi 81936884 81 QSDVNVDNGHFVNIELE 97
gi 82053560 81 QSAVDVDDGHFVNIELE 97
http://www.ncbi.nlm.nih.gov/Structure/cdd/cddsrv.cgi?uid=109647
Vertaan Tukholmalaiseen virukseen , sen M2 proteiiniin.
1 mslltevetp triewecrcs dssdplviaa niigilhlil witdrlffkc iyrrfkyglk
61 rgpstegvpe smreeyqqeq qsavdvddgh fvniele
Tämä on kuin kolmas versio ja sekamuoto kaikista kolmesta.
1..97
/organism="Influenza A virus (A/Stockholm/36/2009(H1N1))"
/strain="A/Stockholm/36/2009"
/serotype="H1N1"
/host="Homo sapiens; gender: F; age: 50"
/db_xref="taxon:657983"
/segment="7"
/country="Sweden: Stockholms Lan"
/collection_date="29-May-2009"
/note="in country transmission via family member returning
from USA"
Protein 1..97
/product="matrix protein 2"
Region 1..97
/region_name="Flu_M2"
/note="Influenza Matrix protein (M2); pfam00599"
/db_xref="CDD:109647"
CDS 1..97
/gene="M2"
/coded_by="join(GQ365365.1:33..58,GQ365365.1:747..1014)"
ORIGIN
1 mslltevetp triewecrcs dssdplviaa niigilhlil witdrlffkc iyrrfkyglk
61 rgpstegvpe smreeyqqeq qsavdvddgh fvniele
JOHTOPÄÄTÖKSENI.
M2 muuntelee suhteellisen runsaasti.
Geeni 7 tuote.
Norjalaisen viruksen M1 proteiini: Sen C-terminaalinen jakso( samanlainen kuin esim katalonialaisen)
/collection_date="08-Sep-2009"
Protein 1..252
/product="matrix protein 1"
Region 2..157
/region_name="Flu_M1"
/note="Influenza Matrix protein (M1); pfam00598"
/db_xref="CDD:109646"
Region 158..252
/region_name="Flu_M1_C"
/note="Influenza Matrix protein (M1) C-terminal domain;
smart00759"
/db_xref="CDD:128998"
CDS 1..252
/gene="M1"
/coded_by="CY052001.1:11..769"
ORIGIN
1 mslltevety vlsiipsgpl kaeiaqrles vfagkntdle almewlktrp ilspltkgil
61 gfvftltvps erglqrrrfv qnalngngdp nnmdravkly kklkreitfh gakevslsys
121 tgalascmgl iynrmgtvtt eaafglvcat ceqiadsqhr shrq//matttn plirhenrmv
181 lasttakame qmagsseqaa eamevanqtr qmvhamrtig thpsssaglk ddllenlqay
241 qkrmgvqmqr fk
Norjalaisen viruksen M2 proteiinista annetaan kaksi hieman poikkeavaa versiota.
//
10 20 30 40 50 60 70 80
....*....|....*....|....*....|....*....|....*....|....*....|....*....|....*....|
gi 81936884 1 MSLLTEVETPIKSGWECRCNDSSDLLVAIASITGILHLILWIFDRLFFKCAYRRFRHGLKRGPSTGGIPESMREEYRQEQ 80
gi 82053560 1 MSLLTEVETPTRNEWECRCSDSSDPLVVAASIIGILHLILWILDRLFFKCIYRRLKYGLKRGPATAGVPESMREEYRQEQ 80
90
....*....|....*..
gi 81936884 81 QSDVNVDNGHFVNIELE 97
gi 82053560 81 QSAVDVDDGHFVNIELE 97
http://www.ncbi.nlm.nih.gov/Structure/cdd/cddsrv.cgi?uid=109647
Vertaan Tukholmalaiseen virukseen , sen M2 proteiiniin.
1 mslltevetp triewecrcs dssdplviaa niigilhlil witdrlffkc iyrrfkyglk
61 rgpstegvpe smreeyqqeq qsavdvddgh fvniele
Tämä on kuin kolmas versio ja sekamuoto kaikista kolmesta.
1..97
/organism="Influenza A virus (A/Stockholm/36/2009(H1N1))"
/strain="A/Stockholm/36/2009"
/serotype="H1N1"
/host="Homo sapiens; gender: F; age: 50"
/db_xref="taxon:657983"
/segment="7"
/country="Sweden: Stockholms Lan"
/collection_date="29-May-2009"
/note="in country transmission via family member returning
from USA"
Protein 1..97
/product="matrix protein 2"
Region 1..97
/region_name="Flu_M2"
/note="Influenza Matrix protein (M2); pfam00599"
/db_xref="CDD:109647"
CDS 1..97
/gene="M2"
/coded_by="join(GQ365365.1:33..58,GQ365365.1:747..1014)"
ORIGIN
1 mslltevetp triewecrcs dssdplviaa niigilhlil witdrlffkc iyrrfkyglk
61 rgpstegvpe smreeyqqeq qsavdvddgh fvniele
JOHTOPÄÄTÖKSENI.
M2 muuntelee suhteellisen runsaasti.
Nyt tulee vaikea ja sekava artikkeli.
Compare H5N1 and Pandemic H1N1 PB2 aminoacids
November 25, 2009
Tässä vertailen vanhaa A H5N1 lintuinfuenssan PB2 aminohapposekvenssiä nykynorjalaiseen ihmisten pandemisen viruksen PB2 proteiinin aminohappojärjestykseen. Erot ovat hyvin suuria. (PubMed lähde)
source 1..759
/organism="Influenza A virus
(A/Goose/Guangdong/1/96(H5N1))"
/strain="A/Goose/Guangdong/1/96(H5N1)"
/db_xref="taxon:93838"
/segment="segment 1"
Protein 1..759
/product="polymerase"
/calculated_mol_wt=85842
Region 1..759
/region_name="Flu_PB2"
/note="Influenza RNA-dependent RNA polymerase subunit PB2;
pfam00604"
/db_xref="CDD:109652"
ORIGIN LINTUUN tarttunut H5N1
1 merikelrdl msqsrtreil tkttvdhmai ikkytsgrqe knpalrmkwm mamKypitad
61 krimemiper neqgqtlwsk tndagsdrvm vsplavtwwn rngpttstvh ypkvyktyfe
121 kverlkhgtf gpvhfrnqvk irrrvdInpg hadlsakeaq dvimevvfpn evgariltse
181 sqlTitkekk eelqdckiap lmvaymlere lvrktrflpv aggtSsvyie vlhltqgtcw
241 eqmytpggev rnddvdqsli iaarnivrra Tvsadplasl lemchstqig gIrmvdilrq
301 npteeqavdi ckaaMglris ssfsfggftf krtNgssvkK eeevltgnlq tlkiKvhegy
361 eeftmvgrra tailrkatrr liqlivsgrd eqsiaeaiiv amvfsqedcm ikavrgdlnf
421 vnranqrlnp mhqllrhfqk dakvlfqnwg iePidnvmgm igilpdmtps AemslrgVrv
481 skmgvdeyss tervvvsidr flrvrdqQgn vllspeevse tqgTekltit ysssmmwein
541 gpesvlvnty qwiirnweTv kiqwsqdptm lynkmefeSf qslvpkaArs Qysgfvrtlf
601 qqmrdvlgtf dtvqiikllp faaappePsr mqfssltvnv rgsgMrilvr gnspvfnynk
661 atkrltvlgk dagaltedpd egtAgvesav lrgflilgRe dkrygpalsi nelsnlakge
721 kanvliMqgd vvlvmkrkrd Fsiltdsqta tkrirmain
NORJALAINEN ( em artikkelissa mainitun viruksen PB2 aminohapot)
ORIGIN IHMISEEN tarttunut pandeminen virus H1N1
1 merikelrdl msqsrtreil tkttvdhmai ikkytsgrqe knpalrmkwm mamRypitad
61 krimdmiper neqgqtlwsk tndagsdrvm vsplavtwwn rngpttstvh ypkvyktyfe
121 kverlkhgtf gpvhfrnqvk irrrvdTnpg hadlsakeaq dvimevvfpn evgariltse
181 sqlAitkekk eelqdckiap lmvaymlere lvrktrflpv aggtGsvyie vlhltqgtcw
241 eqmytpggev rnddvdqsli iaarnivrra Avsadplasl lemchstqig gVrmvdilrq
301 npteeqavdi ckaaIglris ssfsfggftf krtSgssvkN eeevltgnlq tlkiRvhegy
361 eeftmvgrra tailrkatrr liqlivsgrd eqsiaeaiiv amvfsqedcm ikavrgdlnf
421 vnranqrlnp mhqllrhfqk dakvlfqnwg ieSidnvmgm igilpdmtps TemslrgIrv
481 skmgvdeyss tervvvsidr flrvrdqRgn vllspeevse tqgXekltit ysssmmwein
541 gpesvlvnty qwiirnweIv kiqwsqdptm lynkmefePf qslvpkaTrs Rysgfvrtlf
601 qqmrdvlgtf dtvqiikllp faaappeQsr mqfssltvnv rgsgLrilvr gnspvfnynk
661 atkrltvlgk dagaltedpd egtSgvesav lrgflilgKe dkrygpalsi nelsnlakge
721 kanvliGqgd vvlvmkrkrd Ssiltdsqta tkrirmain
Kovin paljon on eroja lintuinfluenssan H5N1 PB2 ja ihmisiin tuleen pandemisen H1N1 PB2 proteiinin aminohappojärjestyksen kesken. Havaitsen näin yhden kerran katsomalla , että
linnusta ihmisen influenssavirukseen on tapahtunut seuraavat mutaatioaskeleet norjalaiseen virukseen nähden:
K54R, (I) isoleusiini147 T, S125G, T271A, (I)isoleusiini292V,
M 315 isoleusiini(I), N 334S, K340N, K355R, P453S, A471T, V477(I),
Q508R, T534X , T559(I), S579P, A588T, Q591R, P628 Q, M655 L,
A684 S, R699 K, M727 G, F741 S.
627 on molemmissa jo E. joten tämä kuvattu lintuvirus on sen osalta jo lajirajan ylittävässä muodossaan.
Aika paljon eroja on kuitenkin tuon vanhan lintuviruksen AH5 N1 ja ihmisissä liikkuvan pandemisen sikainfluenssan kesken.
http://labvirus.wordpress.com/2009/09/05/co-transmission-of-h5n1-with-pandemic-h1n1-in-egypt/
Tarkemmin PB2 polymeraasialayksikön rakenteesta
PB2 polymerase subunit. Aminoacid sequences. November 25, 2009
Vertaan nyt NORJASSA olevaa virusta ja keväällä HOUSTONISSA ollutta pandemista virusta H1N1(2009). Löydän 2 eroavaisuutta niiden PB2-proteiinin kesken.
Influenza A virus (A/Norway/3364-2/2009(H1N1))”
Protein 1..>759
/product="polymerase PB2"
CDS 1..759
/gene="PB2"
/coded_by="CY051995.1:12..>2289"
ORIGIN
1 merikelrdl msqsrtreil tkttvdhmai ikkytsgrqe knpalrmkwm mamrypitad
61 krimdmiper neqgqtlwsk tndagsdrvm vsplavtwwn rngpttstvh ypkvyktyfe
121 kverlkhgtf gpvhfrnqvk irrrvdtnpg hadlsakeaq dvimevvfpn evgariltse
181 sqlaitkekk eelqdckiap lmvaymlere lvrktrflpv aggtgsvyie vlhltqgtcw
241 eqmytpggev rnddvdqsli iaarnivrra avsadplasl lemchstqig gvrmvdilrq
301 npteeqavdi ckaaiglris ssfsfggftf krtsgssvkN eeevltgnlq tlkirvhegy
361 eeftmvgrra tailrkatrr liqlivsgrd eqsiaeaiiv amvfsqedcm ikavrgdlnf
421 vnranqrlnp mhqllrhfqk dakvlfqnwg iesidnvmgm igilpdmtps temslrgirv
481 skmgvdeyss tervvvsidr flrvrdqrgn vllspeevse tqgXekltit ysssmmwein
541 gpesvlvnty qwiirnweiv kiqwsqdptm lynkmefepf qslvpkatrs rysgfvrtlf
601 qqmrdvlgtf dtvqiikllp faaappeqsr mqfssltvnv rgsglrilvr gnspvfnynk
661 atkrltvlgk dagaltedpd egtsgvesav lrgflilgke dkrygpalsi nelsnlakge
721 kanvligqgd vvlvmkrkrd ssiltdsqta tkrirmain
Vertaan tätä johonkin varhaiseen amerikkalaiseen Houstonin virukseen.
Influenza A virus (A/Houston/1OS/2009(H1N1))"
13-May-2009"
Protein 1..759
/product="polymerase PB2"
Region 1..759
/region_name="Flu_PB2"
/note="Influenza RNA-dependent RNA polymerase subunit
ORIGIN
1 merikelrdl msqsrtreil tkttvdhmai ikkytsgrqe knpalrmkwm mamrypitad
61 krimdmiper neqgqtlwsk tndagsdrvm vsplavtwwn rngpttstvh ypkvyktyfe
121 kverlkhgtf gpvhfrnqvk irrrvdtnpg hadlsakeaq dvimevvfpn evgariltse
181 sqlaitkekk eelqdckiap lmvaymlere lvrktrflpv aggtgsvyie vlhltqgtcw
241 eqmytpggev rnddvdqsli iaarnivrra avsadplasl lemchstqig gvrmvdilrq
301 npteeqavdi ckaaiglris ssfsfggftf krtsgssvkK eeevltgnlq tlkirvhegy
361 eeftmvgrra tailrkatrr liqlivsgrd eqsiaeaiiv amvfsqedcm ikavrgdlnf
421 vnranqrlnp mhqllrhfqk dakvlfqnwg iesidnvmgm igilpdmtps temslrgirv
481 skmgvdeyss tervvvsidr flrvrdqrgn vllspeevse tqgTekltit ysssmmwein
541 gpesvlvnty qwiirnweiv kiqwsqdptm lynkmefepf qslvpkatrs rysgfvrtlf
601 qqmrdvlgtf dtvqiikllp faaappeqsr mqfssltvnv rgsglrilvr gnspvfnynk
661 atkrltvlgk dagaltedpd egtsgvesav lrgflilgke dkrygpalsi nelsnlakge
721 kanvligqgd vvlvmkrkrd ssiltdsqta tkrirmain
Kommenttini:
NORJALAINEN on myöhempi, joten asetan sen aminohapon paikanumeron jälkeen ja Houstonin viruksen aminohapon eteen.
K340N, T524X
Houstonin lysiini (K) 340 on norjalaisessa asparagiini (N).
Norjalaisessa on merkintä X, mikä tarkoitaa että siinä kohtaa voi olla ”mikä tahansa aminohappo”. Houstonilaisessa toukokuussa oli threoniini (T) asemassa 524.
LISÄNÄ VANHEMPI HAVAINTO:
REASSORTEMENT, uudelleen järjstäytyminen on havaittu yleisesti tapahtuvan PB2-proteiinin aminohapossa 627 ja sen on havaittu olevan tärkeä determinantti heterospesifisten kompleksien replikaation tehokkuudessa.
E627 on jo näissä molemmissa pandemisisisa H1N1 viruksissa ( Houston ja Norja)
Aiemmin lintuviruksessa oli lysiini(K) ja nyt näyttää sikainfluensaviruksessa olevan glutamiinihappo(E). Siis K627E on tapahtunut aikanaan ja samalla lajirajan ylitys on virukselle helpottunut nykyiseen tilanteeseen.
Lähteenä ovat PubMed uutisten virus sekvenssit.
Keskiviikko, 25. marraskuuta 2009
Vertaan nyt NORJASSA olevaa virusta ja keväällä HOUSTONISSA ollutta pandemista virusta H1N1(2009). Löydän 2 eroavaisuutta niiden PB2-proteiinin kesken.
Influenza A virus (A/Norway/3364-2/2009(H1N1))”
Protein 1..>759
/product="polymerase PB2"
CDS 1..759
/gene="PB2"
/coded_by="CY051995.1:12..>2289"
ORIGIN
1 merikelrdl msqsrtreil tkttvdhmai ikkytsgrqe knpalrmkwm mamrypitad
61 krimdmiper neqgqtlwsk tndagsdrvm vsplavtwwn rngpttstvh ypkvyktyfe
121 kverlkhgtf gpvhfrnqvk irrrvdtnpg hadlsakeaq dvimevvfpn evgariltse
181 sqlaitkekk eelqdckiap lmvaymlere lvrktrflpv aggtgsvyie vlhltqgtcw
241 eqmytpggev rnddvdqsli iaarnivrra avsadplasl lemchstqig gvrmvdilrq
301 npteeqavdi ckaaiglris ssfsfggftf krtsgssvkN eeevltgnlq tlkirvhegy
361 eeftmvgrra tailrkatrr liqlivsgrd eqsiaeaiiv amvfsqedcm ikavrgdlnf
421 vnranqrlnp mhqllrhfqk dakvlfqnwg iesidnvmgm igilpdmtps temslrgirv
481 skmgvdeyss tervvvsidr flrvrdqrgn vllspeevse tqgXekltit ysssmmwein
541 gpesvlvnty qwiirnweiv kiqwsqdptm lynkmefepf qslvpkatrs rysgfvrtlf
601 qqmrdvlgtf dtvqiikllp faaappeqsr mqfssltvnv rgsglrilvr gnspvfnynk
661 atkrltvlgk dagaltedpd egtsgvesav lrgflilgke dkrygpalsi nelsnlakge
721 kanvligqgd vvlvmkrkrd ssiltdsqta tkrirmain
Vertaan tätä johonkin varhaiseen amerikkalaiseen Houstonin virukseen.
Influenza A virus (A/Houston/1OS/2009(H1N1))"
13-May-2009"
Protein 1..759
/product="polymerase PB2"
Region 1..759
/region_name="Flu_PB2"
/note="Influenza RNA-dependent RNA polymerase subunit
ORIGIN
1 merikelrdl msqsrtreil tkttvdhmai ikkytsgrqe knpalrmkwm mamrypitad
61 krimdmiper neqgqtlwsk tndagsdrvm vsplavtwwn rngpttstvh ypkvyktyfe
121 kverlkhgtf gpvhfrnqvk irrrvdtnpg hadlsakeaq dvimevvfpn evgariltse
181 sqlaitkekk eelqdckiap lmvaymlere lvrktrflpv aggtgsvyie vlhltqgtcw
241 eqmytpggev rnddvdqsli iaarnivrra avsadplasl lemchstqig gvrmvdilrq
301 npteeqavdi ckaaiglris ssfsfggftf krtsgssvkK eeevltgnlq tlkirvhegy
361 eeftmvgrra tailrkatrr liqlivsgrd eqsiaeaiiv amvfsqedcm ikavrgdlnf
421 vnranqrlnp mhqllrhfqk dakvlfqnwg iesidnvmgm igilpdmtps temslrgirv
481 skmgvdeyss tervvvsidr flrvrdqrgn vllspeevse tqgTekltit ysssmmwein
541 gpesvlvnty qwiirnweiv kiqwsqdptm lynkmefepf qslvpkatrs rysgfvrtlf
601 qqmrdvlgtf dtvqiikllp faaappeqsr mqfssltvnv rgsglrilvr gnspvfnynk
661 atkrltvlgk dagaltedpd egtsgvesav lrgflilgke dkrygpalsi nelsnlakge
721 kanvligqgd vvlvmkrkrd ssiltdsqta tkrirmain
Kommenttini:
NORJALAINEN on myöhempi, joten asetan sen aminohapon paikanumeron jälkeen ja Houstonin viruksen aminohapon eteen.
K340N, T524X
Houstonin lysiini (K) 340 on norjalaisessa asparagiini (N).
Norjalaisessa on merkintä X, mikä tarkoitaa että siinä kohtaa voi olla ”mikä tahansa aminohappo”. Houstonilaisessa toukokuussa oli threoniini (T) asemassa 524.
LISÄNÄ VANHEMPI HAVAINTO:
REASSORTEMENT, uudelleen järjstäytyminen on havaittu yleisesti tapahtuvan PB2-proteiinin aminohapossa 627 ja sen on havaittu olevan tärkeä determinantti heterospesifisten kompleksien replikaation tehokkuudessa.
E627 on jo näissä molemmissa pandemisisisa H1N1 viruksissa ( Houston ja Norja)
Aiemmin lintuviruksessa oli lysiini(K) ja nyt näyttää sikainfluensaviruksessa olevan glutamiinihappo(E). Siis K627E on tapahtunut aikanaan ja samalla lajirajan ylitys on virukselle helpottunut nykyiseen tilanteeseen.
Lähteenä ovat PubMed uutisten virus sekvenssit.
Keskiviikko, 25. marraskuuta 2009
Entä polymeraasialayksiköt PB2 ja PB1?
Miten tärkeitä viruksen muuttuvuudessa on RNA-polymeraasin alayksiköt PB2 ja PB1 eli virusgeenituotteet geenistä 1 ja geenistä 2 ja niiden suhteet?
Relevance of PB2 & PB1 reassortements in influenza viruses
November 25, 2009
PB2 ja PB1 kombinaatioista
LÄHDE: Li OT, Chan MC, Leung CS, Chan RW, Guan Y, Nicholls JM, Poon LL. Full factorial analysis of mammalian and avian influenza polymerase subunits suggests a role of an efficient polymerase for virus adaptation. PLoS One. 2009 May 21;4(5):e5658. State Key Laboratory of Emerging Infectious Diseases, Department of Microbiology, The University of Hong Kong, Hong Kong SAR, China.
Kaikista A- influenssaviruksen sisärakenteen geenisegmenteistä (PB2, PB1, PA, NP ja NS) on lintuviruksen PB1 segmentti ainoa, joka aikanaan teki uudelleenjärjestäytymisen ( reassortement) ihmisen H2N2 ja H3N2 pandemisiksi kannoiksi. Tästä ollaan päätelty, että viruksen polymeraasialayksikköjen geenien uudelleen järjestäytyminen lämminveristen ja lintujen influenssavirusten kesken saattaisi vaikuttaa virusten siirtymisissä lajista toiseen.
Tutkijat testasivat oletustaan lintujen H5N1 ja lämminveristen H1N1 virusten PB2, PB1, PA ja NP geeneillä ja niiden toisiinsa suhtautumisella.
Luonnehdittiin kaikki 16 mahdollista virus-ribonukleoproteiinin(vRNP) chimeeristä kombinaatiota lintuviruksesta H5N1 ja lämminveristen viruksesta H1N1.
Sitten osoitettiin että sellainen rekombinantti-vRNP, jossa oli lämminveristen influenssaviruksen (H1N1) PB2 ja lintuviruksen (H5N1)PB1, omasi vahvimman polymeraasiaktiivisuuden ihmissoluissa kaikissa tutkituissa lämpötiloissa.
Lisäksi tällä mainitulla tavalla spesifiset PB2&PB1 kombinaatiot omaavat virukset saattoivat kasvaa tehokkaasti soluviljelmissä, varsinkin kuumissa lämpötiloissa (, ilmasto, kuume!). Nämä virukset olivat vahvoja proinflammatoristen sytokiinien ja kemokiinien indusoijia ihmisen primääreissä makrofageissa ja pneumosyyteissä.
Nämä virukset, joissa oli juuri tämä spesifinen PB2-PB1 kombinaatio, pystyivät myös kehkeyttämään adaptatiivisia mutaatioita ( sopeutumisia) uuteen tilanteeseen.
Näistä tuloksista voidaan päätellä, että juuri viruksen polymeraasin aktiivisuus saattaa olla ihmiskunnan terveyden suhteen relevantti tekijä influenssan syntymisessä.
Relevance of PB2 & PB1 reassortements in influenza viruses
November 25, 2009
PB2 ja PB1 kombinaatioista
LÄHDE: Li OT, Chan MC, Leung CS, Chan RW, Guan Y, Nicholls JM, Poon LL. Full factorial analysis of mammalian and avian influenza polymerase subunits suggests a role of an efficient polymerase for virus adaptation. PLoS One. 2009 May 21;4(5):e5658. State Key Laboratory of Emerging Infectious Diseases, Department of Microbiology, The University of Hong Kong, Hong Kong SAR, China.
Kaikista A- influenssaviruksen sisärakenteen geenisegmenteistä (PB2, PB1, PA, NP ja NS) on lintuviruksen PB1 segmentti ainoa, joka aikanaan teki uudelleenjärjestäytymisen ( reassortement) ihmisen H2N2 ja H3N2 pandemisiksi kannoiksi. Tästä ollaan päätelty, että viruksen polymeraasialayksikköjen geenien uudelleen järjestäytyminen lämminveristen ja lintujen influenssavirusten kesken saattaisi vaikuttaa virusten siirtymisissä lajista toiseen.
Tutkijat testasivat oletustaan lintujen H5N1 ja lämminveristen H1N1 virusten PB2, PB1, PA ja NP geeneillä ja niiden toisiinsa suhtautumisella.
Luonnehdittiin kaikki 16 mahdollista virus-ribonukleoproteiinin(vRNP) chimeeristä kombinaatiota lintuviruksesta H5N1 ja lämminveristen viruksesta H1N1.
Sitten osoitettiin että sellainen rekombinantti-vRNP, jossa oli lämminveristen influenssaviruksen (H1N1) PB2 ja lintuviruksen (H5N1)PB1, omasi vahvimman polymeraasiaktiivisuuden ihmissoluissa kaikissa tutkituissa lämpötiloissa.
Lisäksi tällä mainitulla tavalla spesifiset PB2&PB1 kombinaatiot omaavat virukset saattoivat kasvaa tehokkaasti soluviljelmissä, varsinkin kuumissa lämpötiloissa (, ilmasto, kuume!). Nämä virukset olivat vahvoja proinflammatoristen sytokiinien ja kemokiinien indusoijia ihmisen primääreissä makrofageissa ja pneumosyyteissä.
Nämä virukset, joissa oli juuri tämä spesifinen PB2-PB1 kombinaatio, pystyivät myös kehkeyttämään adaptatiivisia mutaatioita ( sopeutumisia) uuteen tilanteeseen.
Näistä tuloksista voidaan päätellä, että juuri viruksen polymeraasin aktiivisuus saattaa olla ihmiskunnan terveyden suhteen relevantti tekijä influenssan syntymisessä.
Muuttuuko PA alayksikkö( Geeni 3. tuote?)
Polymeraasi PA. Muuttuuko tämä helposti vai ei?
Polymerase (PA) of A H1N1 (gene 3): No changes?
November 29, 2009
Norjan uusimmassa A H1N1 viruksessa on PA( geni 3) muuttumaton verrataessa
19 huhtikuuta eristettyyn meksikolaiseen virukseen.
Presiis sama aminohapposekvenssi. (PubMed lähteestä otetut sekvenssit)
/country="Mexico"
/collection_date="19-Apr-2009"
Protein 1..716
/product="polymerase PA"
Region 21..716
/region_name="Flu_PA"
/note="Influenza RNA-dependent RNA polymerase subunit PA;
pfam00603"
/db_xref="CDD:109651"
CDS 1..716
/gene="PA"
/coded_by="GQ379813.1:1..2151"
ORIGIN
1 medfvrqcfn pmivelaeka mkeygedpki etnkfaaict hlevcfmysd fhfiderges
61 iivesgdpna llkhrfeiie grdrimawtv vnsicnttgv ekpkflpdly dykenrfiei
121 gvtrrevhiy ylekankiks ekthihifsf tgeematkad ytldeesrar iktrlftirq
181 emasrslwds frqsergeet ieekfeitgt mrkladqslp pnfsslenfr ayvdgfepng
241 ciegklsqms kevnakiepf lrttprplrl pdgplchqrs kfllmdalkl siedpshege
301 giplydaikc mktffgwkep nivkphekgi npnylmawkq vlaelqdien eekiprtknm
361 krtsqlkwal genmapekvd fddckdvgdl kqydsdepep rslaswvqne fnkaceltds
421 swieldeige dvapiehias mrrnyftaev shcrateyim kgvyintall nascaamddf
481 qlipmiskcr tkegrrktnl ygfiikgrsh lrndtdvvnf vsmefsltdp rlephkweky
541 cvleigdmll rtaigqvsrp mflyvrtngt skikmkwgme mrrcllqslq qiesmieaes
601 svkekdmtke ffenksetwp igesprgvee gsigkvcrtl laksvfnsly aspqlegfsa
661 esrklllivq alrdnlepgt fdlgglyeai eeclindpwv llnaswfnsf lthalk
Polymerase (PA) of A H1N1 (gene 3): No changes?
November 29, 2009
Norjan uusimmassa A H1N1 viruksessa on PA( geni 3) muuttumaton verrataessa
19 huhtikuuta eristettyyn meksikolaiseen virukseen.
Presiis sama aminohapposekvenssi. (PubMed lähteestä otetut sekvenssit)
/country="Mexico"
/collection_date="19-Apr-2009"
Protein 1..716
/product="polymerase PA"
Region 21..716
/region_name="Flu_PA"
/note="Influenza RNA-dependent RNA polymerase subunit PA;
pfam00603"
/db_xref="CDD:109651"
CDS 1..716
/gene="PA"
/coded_by="GQ379813.1:1..2151"
ORIGIN
1 medfvrqcfn pmivelaeka mkeygedpki etnkfaaict hlevcfmysd fhfiderges
61 iivesgdpna llkhrfeiie grdrimawtv vnsicnttgv ekpkflpdly dykenrfiei
121 gvtrrevhiy ylekankiks ekthihifsf tgeematkad ytldeesrar iktrlftirq
181 emasrslwds frqsergeet ieekfeitgt mrkladqslp pnfsslenfr ayvdgfepng
241 ciegklsqms kevnakiepf lrttprplrl pdgplchqrs kfllmdalkl siedpshege
301 giplydaikc mktffgwkep nivkphekgi npnylmawkq vlaelqdien eekiprtknm
361 krtsqlkwal genmapekvd fddckdvgdl kqydsdepep rslaswvqne fnkaceltds
421 swieldeige dvapiehias mrrnyftaev shcrateyim kgvyintall nascaamddf
481 qlipmiskcr tkegrrktnl ygfiikgrsh lrndtdvvnf vsmefsltdp rlephkweky
541 cvleigdmll rtaigqvsrp mflyvrtngt skikmkwgme mrrcllqslq qiesmieaes
601 svkekdmtke ffenksetwp igesprgvee gsigkvcrtl laksvfnsly aspqlegfsa
661 esrklllivq alrdnlepgt fdlgglyeai eeclindpwv llnaswfnsf lthalk
Edelleen PA alayksikön merkityksestä A H5N1 viruksessa
Jotta voi arvioida geeni 3:n roolin käytännössä, pitää katsoa aiempia tutkimuksia PA proteiinin merkityksestä.
LÄHDE: Kashiwagi T, Leung BW, Deng T, Chen H, Brownlee GG.The N-terminal region of the PA subunit of the RNA polymerase of influenza A/HongKong/156/97 (H5N1) influences promoter binding.PLoS One. 2009;4(5):e5473. Epub 2009 May 7. Oxford, Oxford, UK.
N-terminaalinen alue PA alayksikössä eräitten yllämainittujen A-influenssavirusten RNA-polymeraasissa vaikutti promoottoriin sitoutumiseen.
TAUSTA: Influenssaviruksen RNA-polymeraasi on heterotrimeerinen kompleksi, jossa on alayksiköt PB1, PB2 ja PA. Ne tekevät yhteistyötä, saadakseen ensin transkriboitua viruksen RNA-mallisen koodin ja sitten replikoitua virusgenomia. Ne vastaavat geneettisestä puolesta viruksen lisääntymistoiminnassa.
Aiemmin on pystytty osoittamaan, että influenssan PA- alayksikön N-terminaalinen alue eräässä viruksessa A/WSN/33 (H1N1) oli mukana promoottoriin sitoutumiasiassa.Tutkijat halusivat laajentaa tätä näkemystä ja ottivat seuraavia viruskantoja tutkimuksiinsa:
A/HongKong/156/97 (H5N1) ( Ihmisestä eristetty H5N1 vuonna 1997)
A/Vietnam/1194/04 (H5N1) ( Ihmisestä eristetty H5N1 vuonna 2004).
He havaitsivat että molemmissa H5N1 viruskannoissa oli vahvemmin aktiivi polymeraasi, kun verrattiin kahteen klassiseen ihmisen influenssaviruskantaan A/WSN/33 (H1N1) ja A/NT/60/68 (H3N2) koeputkiolosuhteissa.
Tämä lisääntynyt polymeraasiaktiivisuus liittyi promoottoriin sitoutumisasiaan.
Pääasiallinen merkitsijä tässä asiassa oli PA alayksikön N-terminaalinen alue, mistä johtuen promoottiriaktiivisuus lisääntyi.
JOHTOPÄÄTÖS, ASIAN MERKITSEVYYS: Tutkijat ovat sitä mieltä että nykyisten A H5N1 kantojen PA-alayksikön N- terminaali vaikuttaa promoottoriin sitoutumiseen ja he pohtivat mikä tämän merkitys on kantojen virulenssissa .
Overall we suggest that the N-terminal region of the PA subunit of two recent H5N1 strains can influence promoter binding and we speculate this may be a factor in their virulence.
Kommenttini: Ehkä olisi syytä katsoa lintuviruksen N-terminaalinen jakso PA alayksiköstä ja verrata jaksoa norjalaiseen virukseen, joka on tämä pandeminen A H1N1, sillä sen PA alayksikkö on niitä, jotka johtuvat suoraan lintukannasta sikakantaan ilman humaani-influenssavälimuotoa historiassaan. Jos pikaisesti vertailen PubMed lähteen sekvenssejä A H5N1 puhtaan lintuviruksen segmenteihin, poikkeavuus on melkoinen harppaus. En pysty arvioimaan tällaista koska nyt vasta olen alkanut niitä hieman katsella. Koetan etsiä valmiita PubMed artikkeleita , meta-analyysejä, analyysejä.
LÄHDE: Kashiwagi T, Leung BW, Deng T, Chen H, Brownlee GG.The N-terminal region of the PA subunit of the RNA polymerase of influenza A/HongKong/156/97 (H5N1) influences promoter binding.PLoS One. 2009;4(5):e5473. Epub 2009 May 7. Oxford, Oxford, UK.
N-terminaalinen alue PA alayksikössä eräitten yllämainittujen A-influenssavirusten RNA-polymeraasissa vaikutti promoottoriin sitoutumiseen.
TAUSTA: Influenssaviruksen RNA-polymeraasi on heterotrimeerinen kompleksi, jossa on alayksiköt PB1, PB2 ja PA. Ne tekevät yhteistyötä, saadakseen ensin transkriboitua viruksen RNA-mallisen koodin ja sitten replikoitua virusgenomia. Ne vastaavat geneettisestä puolesta viruksen lisääntymistoiminnassa.
Aiemmin on pystytty osoittamaan, että influenssan PA- alayksikön N-terminaalinen alue eräässä viruksessa A/WSN/33 (H1N1) oli mukana promoottoriin sitoutumiasiassa.Tutkijat halusivat laajentaa tätä näkemystä ja ottivat seuraavia viruskantoja tutkimuksiinsa:
A/HongKong/156/97 (H5N1) ( Ihmisestä eristetty H5N1 vuonna 1997)
A/Vietnam/1194/04 (H5N1) ( Ihmisestä eristetty H5N1 vuonna 2004).
He havaitsivat että molemmissa H5N1 viruskannoissa oli vahvemmin aktiivi polymeraasi, kun verrattiin kahteen klassiseen ihmisen influenssaviruskantaan A/WSN/33 (H1N1) ja A/NT/60/68 (H3N2) koeputkiolosuhteissa.
Tämä lisääntynyt polymeraasiaktiivisuus liittyi promoottoriin sitoutumisasiaan.
Pääasiallinen merkitsijä tässä asiassa oli PA alayksikön N-terminaalinen alue, mistä johtuen promoottiriaktiivisuus lisääntyi.
JOHTOPÄÄTÖS, ASIAN MERKITSEVYYS: Tutkijat ovat sitä mieltä että nykyisten A H5N1 kantojen PA-alayksikön N- terminaali vaikuttaa promoottoriin sitoutumiseen ja he pohtivat mikä tämän merkitys on kantojen virulenssissa .
Overall we suggest that the N-terminal region of the PA subunit of two recent H5N1 strains can influence promoter binding and we speculate this may be a factor in their virulence.
Kommenttini: Ehkä olisi syytä katsoa lintuviruksen N-terminaalinen jakso PA alayksiköstä ja verrata jaksoa norjalaiseen virukseen, joka on tämä pandeminen A H1N1, sillä sen PA alayksikkö on niitä, jotka johtuvat suoraan lintukannasta sikakantaan ilman humaani-influenssavälimuotoa historiassaan. Jos pikaisesti vertailen PubMed lähteen sekvenssejä A H5N1 puhtaan lintuviruksen segmenteihin, poikkeavuus on melkoinen harppaus. En pysty arvioimaan tällaista koska nyt vasta olen alkanut niitä hieman katsella. Koetan etsiä valmiita PubMed artikkeleita , meta-analyysejä, analyysejä.
RNA polymeraasi PA alayksikkö( Geeni 3)
Influenssavirusken RNA-polymeraasin yksi alayksikkö, PA ja en olennaiset toiminnot. Influenzavirus RNA polymerase subunit PA, N-terminal
Marraskuu 29. November 29, 2009
LÄHDE: Koyu Hara,† Florian I. et al. Amino Acid Residues in the N-Terminal Region of the PA Subunit of Influenza A Virus RNA Polymerase Play a Critical Role in Protein Stability, Endonuclease Activity, Cap Binding, and Virion RNA Promoter Binding University of Oxford, Oxford, United Kingdom May 23, 2006.
Tutkijat selvittivät tässä PA alayksikön N-terminaalisen jakson merkitystä virulenssissa. He käyttivät puhdistettua rekombinanttia influenssa virusta A/PR/8/34 PA. Sen N-terminaali muodosti 25 kDa domaanin. Sitten tutkijat johtivat tälle evolutionaalisesti ottaen hyvin konservatiiviselle alueelle pistemutaatioita alaniinista. Ne olivat merkitseviä viruksen virulenssille. Jos alaniinimutaatioita asettui L109- F117 jaksolle, siitä seurasi PA proteiinin hajominen, silppuroituminen, millä oli tietysti seuraamuksena polymeraasin huonompi aktiivisuus.
Most alanine-scanning mutations between residues L109 and F117 caused PA degradation, mediated by a proteasome-ubiquitin pathway, and as a consequence interfered with polymerase activity.
Tutkijat selvittivät kolme eri PA:proteiinin pistemutaatiota alaniinin(A) asettamisella virusproteiinin sekvenssiin tiettyihin kohtiin. Lysiini(K) 102, aspartaatti(D) 108 ja lysiini (K)134 muutettiin alaniiniksi (A).
K102A, D108A ja K134A.
(VRT. Allaoleva PA rakenne on norjalaisesta viruksesta. HUOM. Siinä ei ole mitään alaniinimutaatioita noissa kohdissa tietystikään.. Se on evolutionaalisesti stabiili ja siis varmaan "hyvin toimivaa laatua" viruksen arsenaalissa.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1563815/?tool=pmcentrez&report=abstract
="Norway: Akershus" collection_date="08-Sep-2009"
Protein 1..716 /product="polymerase PA"
Numerointi alkaa N- terminaaliseta päädystä:
........1 medfvrqcfn pmivelaeka mkeygedpki etnkfaaict hlevcfmysd fhfiderges
......61 iivesgdpna llkhrfeiie grdrimawtv vnsicnttgv ekpkflpdly dykenrfiei
....121 gvtrrevhiy ylekankiks ekthihifsf tgeematkad ytldeesrar iktrlftirq
....181 emasrslwds frqsergeet ieekfeitgt mrkladqslp pnfsslenfr ayvdgfepng
....241 ciegklsqms kevnakiepf lrttprplrl pdgplchqrs kfllmdalkl siedpshege
....301 giplydaikc mktffgwkep nivkphekgi npnylmawkq vlaelqdien eekiprtknm
....361 krtsqlkwal genmapekvd fddckdvgdl kqydsdepep rslaswvqne fnkaceltds
...421 swieldeige dvapiehias mrrnyftaev shcrateyim kgvyintall nascaamddf
..481 qlipmiskcr tkegrrktnl ygfiikgrsh lrndtdvvnf vsmefsltdp rlephkweky
..541 cvleigdmll rtaigqvsrp mflyvrtngt skikmkwgme mrrcllqslq qiesmieaes
,601 svkekdmtke ffenksetwp igesprgvee gsigkvcrtl laksvfnsly aspqlegfsa
.661 esrklllivq alrdnlepgt fdlgglyeai eeclindpwv llnaswfnsf lthalk
Sekvenssi päättyy C-terminaaliseen osaan.
Huom. Virus on aikamoinen proteiinin kehrääjä ja ottaa tämän valkuaisen ihmiskehosta.
1. Norjalaisessa A H1N1 viruksen PA proteiinissa on konservatiivinen jakso
109L -117F. Siinä ei ole alaniinimutaatioita (A).
Most alanine-scanning mutations between residues L109 and F117 caused PA degradation, mediated by a proteasome-ubiquitin pathway, and as a consequence interfered with polymerase activity.
Jos tässä jaksossa olisi alaniinimutaatioita, PA- peptide alkaisi hajota ubikitiinitietä silppuroituen.
2. K102 A mutaatiota EI ole "kunnon virusproteiinissa" , vaan jäljellä norjalaisessa on K102
Tällainen mutaatio, jos se esiintyisi, vaikuttaisi transkription ja replikaation vähenemistä viruksessa keho-olosuhteissa, in vivo. K102A mutaatio johtaisi sekä cap jaksoon sitoutumisen että viruksen RNA-promoottoriin sitoutumisen aktiivisuuden vähenemistä ja tästä seuraisi estymää sekä virustranskriptioon että replikaatioon.
3. D108A mutaatiota ei ole norjalaisessa , vaan siinä on jäljellä D108.
4. K134A-mutaatiota ei ole norjalaisessa, vaan K134 havaitaan.
Nämä kaksi jälkimmäistä mainittua (tiedemiesten kokeellisesti tekemää) mutaatiota estäisivät selektiivisesti transkriptiota. Molemmat mutaatiot estivät täydellisesti endonukleaasiaktiivisuuden in vitro, mikä selittää selektiivisen transkriptiodefektin.
5. Näistä löydöistä voidaan päätellä, että viruksen PA-proteiinin N-terminaalisessa jaksossa on moninaisia polymeraasifunktioita, joista mainitaan proteiinin stabiilius, endonukleaasiaktiivisuus, Cap-jaksoon kiinnittyminen ja promoottoriin sitoutuminen.
These results suggest that the N-terminal region of PA is involved in multiple functions of the polymerase, including protein stability, endonuclease activity, cap binding, and promoter binding.
Kommenttini:
Siis näissä strategisissa kohdissa ei meksikolaisessa eikä norjalaisessa viruksssa ollut mitään mutaatioita, "viruksen vikaa" , joka olisi heikentänyt viruksen sisätekijöitten virulenssia.
Ei tunneta lääkettä, millä nykyään voitaisiin tähän PA proteiinin N-terminaaliin vaikuttaa ja heikentää virusta. Mutta kuten huomataan, tämä kohta PA-proteiinissa on todella kiinnostava ja terapeuttisen strategian luomisen haaste. Silloin kun keino keksitään, ollaan siirrytty johonkin uuteen menelmään influenssan hoidossa.
Marraskuu 29. November 29, 2009
LÄHDE: Koyu Hara,† Florian I. et al. Amino Acid Residues in the N-Terminal Region of the PA Subunit of Influenza A Virus RNA Polymerase Play a Critical Role in Protein Stability, Endonuclease Activity, Cap Binding, and Virion RNA Promoter Binding University of Oxford, Oxford, United Kingdom May 23, 2006.
Tutkijat selvittivät tässä PA alayksikön N-terminaalisen jakson merkitystä virulenssissa. He käyttivät puhdistettua rekombinanttia influenssa virusta A/PR/8/34 PA. Sen N-terminaali muodosti 25 kDa domaanin. Sitten tutkijat johtivat tälle evolutionaalisesti ottaen hyvin konservatiiviselle alueelle pistemutaatioita alaniinista. Ne olivat merkitseviä viruksen virulenssille. Jos alaniinimutaatioita asettui L109- F117 jaksolle, siitä seurasi PA proteiinin hajominen, silppuroituminen, millä oli tietysti seuraamuksena polymeraasin huonompi aktiivisuus.
Most alanine-scanning mutations between residues L109 and F117 caused PA degradation, mediated by a proteasome-ubiquitin pathway, and as a consequence interfered with polymerase activity.
Tutkijat selvittivät kolme eri PA:proteiinin pistemutaatiota alaniinin(A) asettamisella virusproteiinin sekvenssiin tiettyihin kohtiin. Lysiini(K) 102, aspartaatti(D) 108 ja lysiini (K)134 muutettiin alaniiniksi (A).
K102A, D108A ja K134A.
(VRT. Allaoleva PA rakenne on norjalaisesta viruksesta. HUOM. Siinä ei ole mitään alaniinimutaatioita noissa kohdissa tietystikään.. Se on evolutionaalisesti stabiili ja siis varmaan "hyvin toimivaa laatua" viruksen arsenaalissa.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1563815/?tool=pmcentrez&report=abstract
="Norway: Akershus" collection_date="08-Sep-2009"
Protein 1..716 /product="polymerase PA"
Numerointi alkaa N- terminaaliseta päädystä:
........1 medfvrqcfn pmivelaeka mkeygedpki etnkfaaict hlevcfmysd fhfiderges
......61 iivesgdpna llkhrfeiie grdrimawtv vnsicnttgv ekpkflpdly dykenrfiei
....121 gvtrrevhiy ylekankiks ekthihifsf tgeematkad ytldeesrar iktrlftirq
....181 emasrslwds frqsergeet ieekfeitgt mrkladqslp pnfsslenfr ayvdgfepng
....241 ciegklsqms kevnakiepf lrttprplrl pdgplchqrs kfllmdalkl siedpshege
....301 giplydaikc mktffgwkep nivkphekgi npnylmawkq vlaelqdien eekiprtknm
....361 krtsqlkwal genmapekvd fddckdvgdl kqydsdepep rslaswvqne fnkaceltds
...421 swieldeige dvapiehias mrrnyftaev shcrateyim kgvyintall nascaamddf
..481 qlipmiskcr tkegrrktnl ygfiikgrsh lrndtdvvnf vsmefsltdp rlephkweky
..541 cvleigdmll rtaigqvsrp mflyvrtngt skikmkwgme mrrcllqslq qiesmieaes
,601 svkekdmtke ffenksetwp igesprgvee gsigkvcrtl laksvfnsly aspqlegfsa
.661 esrklllivq alrdnlepgt fdlgglyeai eeclindpwv llnaswfnsf lthalk
Sekvenssi päättyy C-terminaaliseen osaan.
Huom. Virus on aikamoinen proteiinin kehrääjä ja ottaa tämän valkuaisen ihmiskehosta.
1. Norjalaisessa A H1N1 viruksen PA proteiinissa on konservatiivinen jakso
109L -117F. Siinä ei ole alaniinimutaatioita (A).
Most alanine-scanning mutations between residues L109 and F117 caused PA degradation, mediated by a proteasome-ubiquitin pathway, and as a consequence interfered with polymerase activity.
Jos tässä jaksossa olisi alaniinimutaatioita, PA- peptide alkaisi hajota ubikitiinitietä silppuroituen.
2. K102 A mutaatiota EI ole "kunnon virusproteiinissa" , vaan jäljellä norjalaisessa on K102
Tällainen mutaatio, jos se esiintyisi, vaikuttaisi transkription ja replikaation vähenemistä viruksessa keho-olosuhteissa, in vivo. K102A mutaatio johtaisi sekä cap jaksoon sitoutumisen että viruksen RNA-promoottoriin sitoutumisen aktiivisuuden vähenemistä ja tästä seuraisi estymää sekä virustranskriptioon että replikaatioon.
3. D108A mutaatiota ei ole norjalaisessa , vaan siinä on jäljellä D108.
4. K134A-mutaatiota ei ole norjalaisessa, vaan K134 havaitaan.
Nämä kaksi jälkimmäistä mainittua (tiedemiesten kokeellisesti tekemää) mutaatiota estäisivät selektiivisesti transkriptiota. Molemmat mutaatiot estivät täydellisesti endonukleaasiaktiivisuuden in vitro, mikä selittää selektiivisen transkriptiodefektin.
5. Näistä löydöistä voidaan päätellä, että viruksen PA-proteiinin N-terminaalisessa jaksossa on moninaisia polymeraasifunktioita, joista mainitaan proteiinin stabiilius, endonukleaasiaktiivisuus, Cap-jaksoon kiinnittyminen ja promoottoriin sitoutuminen.
These results suggest that the N-terminal region of PA is involved in multiple functions of the polymerase, including protein stability, endonuclease activity, cap binding, and promoter binding.
Kommenttini:
Siis näissä strategisissa kohdissa ei meksikolaisessa eikä norjalaisessa viruksssa ollut mitään mutaatioita, "viruksen vikaa" , joka olisi heikentänyt viruksen sisätekijöitten virulenssia.
Ei tunneta lääkettä, millä nykyään voitaisiin tähän PA proteiinin N-terminaaliin vaikuttaa ja heikentää virusta. Mutta kuten huomataan, tämä kohta PA-proteiinissa on todella kiinnostava ja terapeuttisen strategian luomisen haaste. Silloin kun keino keksitään, ollaan siirrytty johonkin uuteen menelmään influenssan hoidossa.
Prenumerera på:
Inlägg (Atom)